
Blog
Chances are that this blog will die at some point. But it’s a risk I’m willing to take. I will write here short notes, mostly, I guess, on scientific articles I’ve read.
On AI and personal convictions (23/07/2026)
One thing that I think the science-and-AI discourse sometimes lacks is people having stronger convictions. I agree scientists should stay open-minded and ready to update their opinions. But one cannot function without convictions, and when we don’t consciously have any, we tend to unconsciously and sometimes uncritically absorb those floating in the air (‘If you live today you breathe in nihilism’, as Flannery O’Connor wrote in a letter many decades ago) - or just be motivated by optimistic or pessimistic biases.
So I think we need convictions. Educated and careful ones, but convictions nonetheless. What John Henry Newman called ‘intellectual ascent’: you study a certain topic, you try to discover the truth or what seems likeliest to be the truth, and then you ‘ascent’ to it with your will, a human faculty distinct from intellect that motivates action. You keep it careful and humble and open-minded but you do believe in certain truths. And you are ready to take responsibility for those convictions: you are ready to have ‘the skin in the game’, you take the risk and are willing to take it.
Many of my convictions are philosophical and ethical in nature. I have a certain view of the importance of human beings, of their capacity for good and truth, of human responsibility; also of the larger scheme of things in which our human pursuit of the good fits in and in which computers don’t fit in other than as tools. All that affects the way I see AI and its place in human work and life (for example, I am strongly against all anthropomorphising attitudes, and I think it should be legally regulated for AI not to mimic the way humans speaks). But I also have some more concrete convictions about what I think computers can and cannot be or do for science in principle, and also even more concrete, if riskier, convictions about where AI in science is likely heading. I’m not claiming I’m suddenly the only person out there who can predict the future, obviously I cannot. But neither am I someone who constantly says ‘for now it’s like this, in two years it might be completely different’ (‘for now AI is not autonomous in science, soon it might be’; ‘for now it is not conscious, soon it might be’, etc.). No.
This post is in part motivated by a depressing interview with a mathematician, the prospective Fields medalist Jacob Tsimerman, where he says, for example, ‘I think there will be a point where AI will be strictly better than humans at all aspects of math: learning, proving, coming up with the problems, aesthetics. It will just be better at everything. And this will come pretty soon’. When asked how soon, he says maybe five years, with big bars of uncertainty. He also says there are many ways in which AI may kill all of us, and that it will be a win if it just does everything but lets us live in peace.
Now this might sound like just staying open-minded about the future and avoiding naive optimism, but obviously if you ascent to these kinds of views as likely, it affects everything you do. And mostly in a gloomy kind of way. I am not saying the author of these words thinks like this necessarily, but I have met many people who find this kind of vision convincing (sometimes against what they would prefer to believe) and very depressing and demotivating at the same time.
So I feel like it is important, for those who have studied the matter sufficiently and are convinced it is not true, to own and act on those convictions. I, for one, don’t believe ‘AI will be better at everything’ in any domain (see footnote). In fact, one thing I do believe in is that, just by the nature of how things work, AI will, as it improves, always tend towards a certain high level of human capability in a given task (but do things at that level much faster, and thus becoming a tool with its own niche: automatising a certain high level). A high, very high level but nonetheless not the highest possible, nor the level one should aim for, not a satisfactory level, leaving a space where humans can and should make all the difference if they want. If this is ‘moving the goalposts’, it’s because what we mean by ‘high level’ or ‘very high level’ will change, but I do believe there is an inherent ceiling relative to human performance.
One of the cases that informs these convictions is AlphaFold, its capabilities and limitations, its reception in our protein structure/biochemistry community. It is astonishing how good it can be at the task of proposing rigid structural models of biological things. Surely rigid models are themselves an approximation, but a useful one, and being able to propose accurate models is a worthwhile task with which AlphaFold can help. What it proposes is very often correct as a rigid model. But it will never be ‘completely and always correct’ (even as a model), of the very nature of things (because it a compressed image of the experimental training set from which it tries to extrapolate etc. etc.), and that is the gap where human intervention, probing things deeper, reflecting from different angles, thinking for years and not hours, testing experimentally etc. makes all the difference. The gap does not seem to get that much smaller with new protein prediction algorithms (e.g. AlphaFold3 vs AlphaFold2), because at some point there is only so much you get out of the data by means of generalising and by not having a real intellect in the loop but only computation (you guessed it, I hold convictions about human intellect being distinct from mechanical computation). And additionally this small but crucial gap between what AlphaFold proposes and the reality, is only small if the person using AlphaFold is trained in protein biochemistry and in the use of this particular software. If the person using it isn’t properly trained, there are additional limitations linked to mis- or overinterpretation, not knowing the common blind spots of the model, not to mention simply not understanding what one looks at and what for.
Now one of the views I strongly hold is that this sort of description is roughly representative of all AI, current and future. Obviously AlphaFold is just a metaphor here. The largest available LLM models that are so good at certain mathematical tasks, for example, are clearly quite different, and one can also perhas use them, if one knows how, to initiate oneself into a given discipline even if one wasn’t trained in it (although at some point there is a human limitation of how fast one learns and how many things one can hold in ones head etc. that a computational advance cannot bypass; a real, hands-on, guided, tacit-learning-aware training is needed most of the time). I’m not saying AI will not be astonishing, transformative etc. - but these basic things will always remain true.
So, in short, the following is the most convincing metaphor for thinking of AI that informs my action: ‘good at tasks until a certain gap, making the gap smaller with time, but always leaving the gap there, and a gap that makes a critical difference and can and must be bridged by human effort, and humans often being able to reach there without using AI to automate things up to the gap in the first place - and perhaps sometimes only being able to reach there if not using AI on the way’. To this I would add the conviction that the way, rather than the destination, is what is often important, and this affects also the attitudes we should have. And finally, we must think on a larger scale, which means in part thinking of our place in the society, of how we as scientists build a relationship with the rest of the society, in dialogue, but also with a defined role for us as a sort of reference point for certain topics, which puts a lot of responsibility on us to work on ourselves (including our morals, our meta-scientific sense of what is true and worthwhile, how various insights fit together etc.) as much as on generating scientific results.
With all this in mind, I think mentoring younger people has never been more important. Also working on self-discipline and on limiting use of computers in certain areas. On thinking through ethical and philosophical topics and being ready to cling to your convictions and be willing to take the risk in their name. Working as a team and as part of a society. Artisanal science that does not use any AI is not a bad idea - definitely not when seen as a choice among others to diversify available attitudes - although perhaps some use can be beneficial or even strongly beneficial overall, when done in a responsible way (this also means thinking of societal and ecological aspects of things, which is difficult with current models).
P. S. ‘I, for one, don’t believe ‘AI will be better at everything’ in any domain (see footnote)’. Aren’t things like chess or GO an exception? The problem with such examples is that human games have artificially imposed unchanging rules and a clear well-defined goal, for which a model can be trained. Perhaps there are parts of maths that are a little bit like this. But most other domains of human activity, when understood sufficiently broadly, are about constantly rediscovering what the goal is and what the rules are.
Metaphors for AI (21/05/2026)
I’ll write again about “AI” - this time in connection with today’s news that an internal OpenAI model solved an interesting mathematical problem in discrete geometry (specifically, it proved that a certain conjecture in discrete geometry is false). I’m of course no expert in real, research-level mathematics, but I’ve tried to follow the progress in the use of “AI” in this field over recent months, and I understand the academic system well enough to know who counts as an authority and so on. And in this case, for example, Timothy Gowers - one of the most respected living mathematicians - says this is a big breakthrough, something that some mathematicians had tried to achieve and never fully managed, something that would have been published in the most prestigious mathematical journals had it been a human achievement. So it’s not something unheard of in human mathematics, but it is nonetheless a technological breakthrough, and probably a sign of further breakthroughs to come - in mathematics and perhaps in other areas of science and life (though one should of course remember that OpenAI surely tried the same trick on hundreds of important mathematical problems and announced the one case where it worked).
And hence this post. The post will be about the metaphors that come to my mind when thinking about so-called “artificial intelligence”, or more broadly, metaphors perhaps useful (or so I think) for the era of so-called “artificial intelligence”. So-called, because “artificial intelligence” is itself a metaphor - and not a particularly good one at that.
Cooking pots from a TV shopping channel. As a first one, perhaps the recent metaphor from a post by a Polish writer Szczepan Twardoch: that AI is like the pots in the 1990s in Poland (during Polish capitalist transition that saw a lot of TV shopping shows popping up) that someone tries to push on you, praising how good they are, how essential. AI as the new TV-shopping pitch - except with a technical, scientific aura - and with real, enormous, though exaggerated, potential. Even the very fact that AI companies have now started focusing specifically on mathematics is probably, to some extent, a marketing move. Fundamental mathematics is a field that is usually underfunded and forgotten, yet highly prestigious in the public imagination - and on top of that, a discipline in which certain things can be proven without experimental verification. Proven, so as to sustain faith in “AI”, to sustain the constant tension, the eternal promise of an “enormous breakthrough” about to arrive. So companies have directed part of their enormous funds and human talent toward mathematics, and they’re getting results. Whether this translates in any way into the value of “AI” in other areas of life is uncertain. And the problems with “AI” - appropriating human knowledge and creativity without paying for it, polluting the environment, lack of transparency, marketing hype around everything, littering the world, stripping human work and creativity of their value - remain.
A shared dinner. This is a metaphor proposed recently by the eminent mathematician Terence Tao. It goes roughly like this. In so-called primitive societies, where food was scarce, the most important work was done - and the greatest respect was enjoyed - by whoever brought the food. But now, when we have plenty of food, bringing a piece of meat of unclear origin to a shared dinner is not a significant contribution to a community meal. What matters more is choosing trusted ingredients, preparing them in a way that makes food digestably and tasty to everyone, presenting food, and dividing it up. So perhaps in an era when a machine easily proves mathematical theorems, we need to reassess what matters in mathematics: what becomes important is not the proving itself, but understanding a new proof and making it easier for others to understand it. Not to mention that a large part - some say the main part - of mathematics is not proving theorems, but posing conjectures, formulating definitions, creating new fields. Similarly in other areas of academia and life: the automation of certain tasks triggers a reassessment of a given field; whatever is easy and plentiful becomes of little value. What becomes key is selection, and whether the new things genuinely advance human understanding of reality and help build a human community of understanding.
The blender. Terence Tao also mentions another metaphor, which I had earlier heard from David Bessis: the metaphor of the blender. The fact that it’s easier for us to digest blended food doesn’t mean we want to blend everything. We don’t, because the “resistance” offered by unblended food matters for taste and health. “AI” makes many things easier and faster - including obtaining information - but in that easing there is also a problem, because we learn, develop, and remember when we encounter “resistance”: when we have to make an effort to find, solve, program, translate, or write something. Hence the need to consciously give up “AI” - at least in certain fields, at least at a certain age, at least during certain periods. I think that as a rule we should give it up whenever we can, and certainly in areas where we want to keep our own abilities and voice.
Botox. This metaphor comes from an essay by Anna Jordan in The Plough. Editing texts with “AI” is like addictive “botox” - it smooths out wrinkles, and once we’ve convinced ourselves (which comes easily) that we must smooth them out, it’s hard to break free. But sometimes - usually - it’s the wrinkles that are interesting. And above all - they’re real.
Classical music. In a sense I envy classical musicians. Unlike chess players, unlike mathematicians now, and perhaps people in other fields of science too - nobody compares them to machines. It’s well known that a machine can produce a pure sound. It’s well known that high-quality recordings capture masterful performances of musical works. But to perform a piece of music live as a human being - that is a task in its own right, one at which, by definition, a human is the best. And a human, if they practice a great deal, can reach an astonishing level, and even if they practice only a little, they will usually reach a level satisfying enough to enrich their own life and the lives of others. I think that very many human tasks - perhaps all of them, to some degree - have at least an element similar to classical music: that is, at least part of the value of everything we do comes from the individual human element present in it. And that is something no automation can, by definition, achieve or surpass.
The library juicer. “AI” companies try hard to present their products as tools that do something on their own. The truth, however, is that these tools resemble more a juicer, which in itself has little value without material to squeeze the juice out of. That material, in the case of “AI,” is human knowledge accumulated over centuries. For example, all of mathematical knowledge - every existing solution to solved problems. This matters. For instance, today’s discovery, instead of “AI solved a problem”, could be called: “existing mathematical knowledge was successfully mechanically extrapolated in a way that allowed a new open problem to be solved”. As a juicer, “AI” is at once astonishing and extremely unreliable. Astonishing, because it finds things quickly - it’s a real, finally working semantic search engine. Astonishing, because in fields such as mathematics, where the origin of an idea is secondary, where logical connections are tight, and where methods of verification exist, it can build very valuable constructions. But also unreliable, because it has a tendency toward errors, distortions, and fabrications. Even in mathematics, much of what “AI” proposes is untrue or a dead end - one has to know how to verify, select, and steer it. In fields such as history, where the origin and details of an idea, and even the way it is phrased, are of fundamental importance, “AI” as a juicer, in a sense, distorts things by definition, and perhaps is altogether useless beyond the basic semantic search application.
The car and the bicycle. It’s no great feat to predict the car. The feat is to predict the traffic jam. The quote is supposedly from the science-fiction writer Frederik Pohl. The fact that “AI” can do many things faster than we do them now is one thing. The full consequences of this technology - including the unforeseen negative ones - are another. Personally, I prefer the bicycle to the car. It’s astonishing how far you can travel using your own legs. You’re healthier. You get more satisfaction. You don’t cause traffic jams. At the same time, I understand that ambulances are cars not bikes. But it is rarely a value in itself to do something as fast as possible. Even cars could drive much faster than they do, but then they would be too fast for people to use safely and beneficially. Producing a million lines of mathematical content per minute - most of them wrong, on top of that - is no great feat. Doing all of it at a pace that moves forward our shared understanding of reality and human intellectual potential - that is an entirely different task, even though mechanical proofs of new theorems can certainly be useful.
Little balls flying around blindly until they find an exit. Ultimately, every computer problem has somewhat the character of blind movement in a cage with an “exit”. This is the case, for example, with programming using “AI”: the system tries and tries until it completes a verifiable programming task. That is something entirely different from human rational construction. The first method can often be very effective, but it isn’t suited to everything. Sometimes it’s hard to fit it into existing structures. And it isn’t very effective when there is no single simple “exit” - no clearly verifiable goal. For instance, for centuries we’ve been arguing about what the goal of literature is - so how could a program be created to fulfill that goal?
AlphaFold. As it happens, in my own field of science, structural biology, we had a significant breakthrough a few years ago brought about by the “AI” of that time - by AlphaFold, which works like a “juicer” of existing biochemical knowledge, making it possible to predict protein structures fairly accurately - something that was previously impossible, and is very important. My field passed quickly through various stages. At first there was something like denial: it’s impossible that this works. Then exaggerated fear or delight: it works 100%, we’ll all lose our jobs, biology is solved, and so on. In the end the situation stabilised. AlphaFold models are extremely useful, but not 100% certain. A whole subfield arose, devoted to optimising them, verifying them, and finding new applications for them. Nobody lost their job, though people had to readjust. But, perhaps most importantly, AlphaFold became so valuable right away precisely because there were plenty of structural biology experts - trained in the era before AlphaFold, when things required more effort - who were able to appreciate it, apply it to their own purposes, judge the quality of the models obtained, and verify them.
A few thoughts on Harvey Lederman’s essay on AI and the meaning of life (19/05/2026)
Someone wrote they found the essay ‘annoying’. It might be the right word to describe my feelings, too. I feel bad about this, because the essay is thoughtful in tone, and the author sounds like - and I’m sure, is - a nice chap, the kind of person you would want to have as a lecturer, or a friend. Also, the essay is personal, sincere; the author makes himself vulnerable, mentions his inner fears, his family. That is why I feel bad about, in part, disliking it. But I feel the author wants there to be a debate.
Perhaps the part I most object to is where he mentions the hypothetical (but presented as likely, at least in a (very) distant future) scenario where, if we love our children, we might feel obliged to hand over parenting to bots and robots that, unlike us, will ‘never snap’, be infinitely patient and understanding etc. I felt a strong objection to that all the more because our third child has just been born three weeks ago, and I’m reminded once again of the indescribable joy of taking care of a young child. Part of this joy is, I think, that you learn to trust yourself, and that you discover you are capable of more care and love than you thought you were. Being afraid of ‘snapping’ is, incidentally, a common OCD symptom developed by a small percentage of young parents, usually the more thoughtful and careful among them, in which, overwhelmed by ‘what could happen’ (but never actually happens for the parents obsessed by it), they live in fear of themselves. Luckily I have been mostly spared that, and I can enjoy the feeling of growing in confidence, despite feeling, in principle, incapable and unworthy. Life is so complex, isn’t it, that sometimes the right thing is to trust - in this case, yourself. How do people hypothesising about parenting bots imagine it will actually happen? Will the robots be better at parenting by definition, or will people do large-scale clinical trials first? I’m a biologist, so one thing I learned is that if you imagine a certain therapeutic thing will work, your imagining is not enough to prove it does - you have to first test it, and then most of the time you discover it doesn’t work that well, and there are many unexpected secondary effects. Similarly, human-robot interaction is a complex domain of its own, and a very unpredictable one at that, as seen, for example, by recent cases of suicides to which interactions with chatbots have apparently contributed. And, if things do become concrete and such hypothetical clinical trials are to be organised, how will certain parents be morally justified in enrolling their children in such trials without first knowing it’s safe? And how long will the trials be conducted to be sure of the effects? What if they take a generation to emerge? And isn’t it an ethical problem if a machine simulates care that presupposes feelings?
These sort of paradoxes - and one could list many, many more - remain unresolved in this essay. Nor is it its purpose to resolve them, and I respect that (although I think that the fact that such things are not considered at all is telling - that the framing here is one based mostly on imagining how things should develop, with little real engagement with messy, unpredictable reality). The purpose of the essay is to reflect on a hypothetical situation, heralded by the arrival of ChatGPT and likely, we are told, arriving at some point in the future, where we have to hand over to artificial intelligence many things we cherish: first intellectual work, but eventually, perhaps, even parenting, presented as a sort of extreme - all that for the sake of a ‘better future’ (that such a future will ultimately be better, the author does not seem to doubt, except for a slight possibility that robots might kill us before such a future materialises).
The response the author proposes to this bitter-sweet situation is grief. Grief like that of his in-laws from a beautiful Italian mountain village, who have to learn to live with the fact that the world is changing, their way of life is going away, people stop growing chestnuts and using the many words for ‘chestnuts’ present in the traditional dialect. These changes one must accept, but at least one can grieve - that is the message of the essay that I got out of it. The grief - this at least no one can take away from us. There is another thing, I forgot, that according to the author, no one can take away from us either. Namely, the task of internal self-fashioning, of creating oneself as a subject. This I’m not that sure about, though. If we might feel obliged to confide our children to bots because they are better at parenting, why not confide ourselves, our internal self-fashioning, to a bot? Why endanger my future self? What if I take a wrong course that distorts my future self instead of making it internally beautiful? By the way, we might give away the task of self-fashioning to bots anyway even if we don’t choose to, because we aren’t as autonomous as we like to think, and even by interacting with bots we do get influenced by them, as many recent observations and studies suggest.
The grief that the author prescribes sounds wise. But it might also be seen as a way of suppressing resistance. In contrast to wise grief, resistance, perhaps even measured, informed resistance, would be seen as futile and immature. I referred to parenting OCD on purpose before, to highlight that our perception of reality - including of what is risky, what should be prevented, which values prevail when values are weighed against each other, particularly when one value is some intrinsic good and the other is harm prevention - is not as straightforward as it seems. Usually there are different levels of response possible, and different values to be reconciled. In democratic societies we negotiate these things - we do so also in the families and communities we live in; we do so inside ourselves, through internal dialogue. There is no mention of any democratic negotiation in the essay, not even as something that would be “good to have” in principle, however unrealistic. The future governed by bots is something that will come and will be good to us in the end - we seem to be told - and if we don’t like losing certain things we cherish now - we are always allowed to grieve.
I would suggest one thing. Perhaps it is useful to look at how we treat non-human communities (animal ecosystems), or even human, endogenous communities - and apply that sort of thinking to ourselves, at least to some extent. We do usually try to protect ways of life, we do try not to interfere too much, not to over-optimise according to our imagined rational view, not to overengineer; to trust, to some extent, in “how things are”. Sure, technology can be introduced, but with a very careful approach, with “conservation” of what exists and is good always in mind; with clinical trials to see if there are unexpected consequences. Given that such thinking is arguably more prominent in Europe than in the US, I wouldn’t be surprised if this kind of approach informed the (perhaps invisible, but real) conservation policy in beautiful Italian villages that still exist, despite the changing world, and which might, who knows, see better future again, given that we can partly study or work through Zoom…
And, lastly, a word regarding whether the future imagined in the essay might actually come to pass. I don’t downplay the capability of AI in certain domains (including, increasingly, in the scientific domain), although I perhaps interpret it differently from some people. Yes, the amassed corpus of human knowledge, including mathematical and scientific knowledge, has proven to be extrapolable, in a complex way, to produce new insights. Patterns, methods, perhaps even local ways of reasoning can be deduced and applied to new problems in a way that is somewhat messy but often very impressive. The main “thing” doing all that is the human corpus of knowledge itself, which turned out to contain more independent potential - independent from new human interventions, that is - than we assumed (but we are still to give enough credit to all the human beings who contributed to it, and should benefit from it and have a say in how it is used). AI is (just) a way of deriving these insights - even if the companies making it seem to want us to forget about the human work that went into it and focus on the tool as a semi-autonomous, quasi-human “agent”. That said, the emergence of AI is a completely new phenomenon with many unpredictable consequences, including the consequences of our (over)reliance on it, and further technological advances - including robotics - are of course on their way. But perhaps - if we don’t just resort to grief, but also act, and, crucially, don’t lose confidence and trust in ourselves, nor become overconfident in this (very messy) technology - we can strive to make our future ever more beautiful. Including in science and academia, and in parenting. Meanwhile, I’m still on paternity leave until the end of May.
A few thoughts on ‘sticky pockets’ for linear motif binding (07/04/2026)
Among other topics, we study SIMs (SUMO-interacting motifs), which are a good example of simple short linear motifs that bind to dedicated pockets on their target proteins. AIM/LIR motifs, which mediate interactions with ATG8 autophagy proteins, are another, similar example - perhaps an even better one. Interestingly, SUMO and ATG8 proteins are evolutionarily related, which raises the possibility that a proto-SUMO/ATG8 protein once bound to proto-SIM/AIM motifs in the distant past, although these kind of things are difficult to know with certainty). What makes such pockets “sticky”? I think the key feature may be their ability to show very low but meaningful affinity even to rudimentary motifs encountered in proteins by chance. In some sense, you can only optimise in evolution what you already have at least in a rudimentary form. So if the binding pocket has weak but appreciable affinity for simple motifs that occur frequently by chance, specific substrates can then optimise those interactions if advantageous. At least that is what I like to think, although this assumes that acquiring the initial rudimentary affinity is the kinetic bottleneck in evolution. Structurally, ‘sticky pockets’ often seem to involve beta augmentation: adding an extra beta strand to an exposed beta sheet. This makes sense, because strand:strand interactions are partly main chain-mediated and thus more loosely dependent on sequence. In the case of ATG8, there is also a deep pocket for an aromatic “hook” - surely another ‘sticky’ feature. But having a pocket for which simple binding motifs emerge readily de novo is a double-edged sword. It gives the system an advantage - you can easily expand an interaction network in evolution. Yet it also creates a vulnerability, as pathogens can hijack those same pockets, and perhaps cancer could too (do we know any such examples, of linear binding motifs created de novo through cancer-associated mutations?). Such features - an exposed beta sheet, a pocket for an aromatic - both attract positive evolution and attract hijacking. You would expect them to exist in those proteins where expanding the interaction network justifies the risk.
Redesigning protein sequence for stabler proteins - intrabodies and other cases (25/03/2026)
Very rare to have two posts on the same day, but this is just to say that, in yesterday’s journal club, we discussed this nice paper on transforming antibodies into intrabodies from Timothy Stasevich et al. The authors put together a pipeline comprising ProteinMPNN to design a stabler protein sequence for the antibody fragment scaffold while keeping the antigen-binding loops unchanged. The idea behind the paper - nicely presented yesterday by Aanchal and Rafael from our team - is that just taking fragments of known antibodies and expressing them in cells usually doesn’t work, as antibodies are optimised for the extracellular, oxidising milieu, but redesigning the scaffold can render them functional. This doesn’t work every single time, but is sufficiently efficient to make the pipeline worthwhile at scale. It reminded me of this paper we discussed some time ago in a journal club as well, where the sequences of membrane proteins were redesigned with a similar pipeline while keeping the business bits unchanged in order to create soluble functional and structural analogues of membrane proteins. And now even more broadly: I guess this could be used in many biological contexts where a function depends on a particular spatial arrangement of amino acids, but the protein is not easy to handle - just like in these two examples, one could keep the key amino acids while redesigning the rest of the sequence to get a stabler protein with a similar structure. An interesting question is why redesigned sequences tend to be stabler than natural ones. I guess that - in addition to these tools being trained mostly on stabler-than-average sequences from the PDB - there is something about natural sequences that usually renders them suboptimally stable, and not only when you take them out of their natural context (as with antibodies that were fragmented and put into cells or membrane proteins taken out of membranes) but even if you keep them in their native setting. I like this paper reminding us that evolutionary optimisation is about balancing conflicting parameters. For natural proteins, structural stability is just one parameter to optimise among many others (including dynamic aspects, which are in a more less direct conflict with structural rigidity), so evolution makes them less stable than an ‘averaged’ synthetic sequence that just maximises structural fit. But it also means that, for therapeutic uses for example, redesigned sequences might often be wanting as they optimise only one among many important aspects of a protein - e.g. I can easily imagine that they render the protein too highly expressed, too difficult to degrade, insensitive to allosteric signals, incapable of some unforseen binding events, too soluble, or even catalytically inactive by lacking required dynamic structural transitions. But for research tools - a redesigned stable, crystallisable, highly expressing sequence folding into a desired structural epitope could be just what you need.
Native holdup for protein:DNA interactions (25/03/2026)
Just finished reading this lovely paper by Boglarka Zambo, Gergő Gógl, Gilles Travé, and collaborators, describing a new technique for quantifying transcription factor:DNA binding without protein purification. We have been interested in transcription factors for some time (see especially this article on ZBTB transcription factors) - and found them very difficult to produce recombinantly in full-length forms - so this technique is super exciting to read about. If you haven’t heard of native holdup assays - developed by Gilles Travé and coworkers, including Gergo and Boglarka - they measure binding between molecules A and B by quantifying how much molecule A is depleted after incubation with immobilised B. It’s basically the opposite of a pulldown, and, by being more specific without the need for washing steps, it’s more quantitative and compatible with transient interactions. You probe the levels of A before and after depletion with mass spec. or any other suitable technique, e.g. western blot. A holdup with a single concentration of immobilised B is in theory enough, but a titration with multiple B concentrations is more accurate. So here the authors extend the native holdup approach to protein:DNA interactions, which requires some finetuning. The beauty: no need to purify your protein - it can be present in cell lysate. A gamechanger for transcription factors, so difficult to purify in full-length forms. You simply measure transcription factor levels in cell lysate after incubation with known concentrations of immobilised annealed DNA oligos. At least on paper, this looks easy and potentially so informative. Look at figure 6 in the paper (panels A and B): GATA1 levels in K562 extract are probed by WB after incubation with increasing concentrations of immobilised cognate or mutated DNAs. High DNA concentrations deplete the TF; with less DNA, more TF remains. You quantify the bands to get binding curves with apparent Kd. You see that one DNA probe (51) binds tighter than the other (4945) - if I remember correctly, the difference is in the number of GATA1 binding motifs. Consistently, mutated versions of the DNA probes deplete less, giving higher Kds. And in the next figure, they repeat the same experiment but with lysates of cells expressing WT and mutants GATA1s to compare their affinities for the same DNA probes. One of these beautiful, elegant papers. And it’s nice to see it in the FEBS Journal! Congratulations to Boglarka, Gergő, and all the authors!
A few more thoughts on LLMs in science (16/02/2026)
Returning to the topic of using frontier LLMs for maths, after the recent 1stproof initiative (real research questions by mathematicians announced to test AI). A few observations:
1) One striking thing is laymen who attempted solving problems without knowing any math, believed their solutions must be true because the models were convinced (even when crosschecking with official solutions) - while they weren’t. They were even more convinced when formalising their solutions in Lean and getting a validation, but formalisations often seemed nonsensical too.
2) Another thing one could see - or perhaps more ‘sense’ - was everybody (the laymen participating in the contest, the audience) hoping experts would look at and comment on the various solutions. But the experts don’t necessarily have time to go through all that, so the things remain unresolved, uncertain. With AI and its potential for research slop, the value of experts who can deeply understand and evaluate things only increases.
3) OpenAI apparently cracked some problems with their ‘internal model’. They announced it immediately and used it for PR. Reactions as expected: AI does maths better than humans etc. etc. But they used human supervision and wouldn’t share transcripts — while 1stproof was meant for unsupervised runs.
4) Frontier LLMs for maths (and other areas of science) seem to work mostly where similar approaches and ideas were used somewhere in the literature but need to be extended or, perhaps, combined. AI as a great semantic search for ideas, but you don’t know if what it proposes makes sense. It takes an expert to ask a question and navigate answers. But it potentially gives you a new way of doing certain aspects of science. This capability is nicely described in this quote from the mathematician Terrence Tao: There has been a really visible increase in capability. It is not pure hype by any means. To me, these advances show there is a complementary way to do mathematics. Humans traditionally work in small groups on hard problems for months, and we will keep doing that. But we can also now set AI to scale: sweep a thousand problems and pick up all the low-hanging fruit. Figure out all the ways to match problems to methods. If there are 20 different techniques, apply them all to 1,000 problems and see which ones can be solved by these methods. This is the capability that is present today.
Overall, AI both impressive and disappointing; the science (maths in this case) both changing profoundly and real human expertise becoming even more valuable; AI companies making real progress but also constantly manipulating reality, distorting the public view of what science is about.
I don’t envy people who start their PhDs now. What they should be trying to do — I think — is to first become confident experts in certain things, and this might involve using as little AI as possible not to become dependent or have their view distorted.
P.S. I know very little about maths and I’m following the current debates purely as a layman, who tries to draw parallels with biochemistry.
Parallels between machine learning in maths and biology? (20/01/2026)
I find the current developments in AI’s use in mathematics interesting (see e.g. some recent posts by Terrence Tao on mathstodon.xyz/@tao, one of which I paste below). It does feel a bit like ‘an AlphaFold moment’ in the sense that suddenly you don’t need hype because there is a real thing happening, with ‘AI’ - or shall we call it machine learning/automation? - providing new possibilities in this particular branch of research (another area where this seems to be happening is some types of coding).
I used to love maths but know little of its professional academic side, so whatever I think of it is surely very naive, so please forgive me. The idea is more to look at it from outside, as a structural biologist.
I’m old enough to remember that prior to AI, computers were already helping to prove maths problems, like with Fermat’s Last Theorem within certain range (prior to a general proof by Andrew Wiles), or a Four Colour problem, or, more recently, the Kepler’s conjecture. But now it feels different, with ‘reasoning LLMs’, some of them maths-specific, some general like ChatGTP 5.2 Pro, single-handedly offering valid proofs to some fairly complex problems, such as those on the Erdős list (although arguably only to the ‘low hanging fruit’ on this long list). For the summary and discussion of AI’s contribution to these problems, which exploded in recent months and, especially, weeks, see this site.
While I very superficially understand all this, what I want to do in this blog is to consider an analogy between what is happening in maths now and what happened to structural biology with the arrival of AlphaFold (AF) a few years back. First of all, AF does not ‘solve structures’ by itself - it is just generalising from the human-made body of experimental structures, similarly to AI’s maths contributions being generalisations from the large body of human mathematical work - the value is more in that work and its potential than in an ability to computationally harvest that. Secondly, AF has not really invalidated any older biochemical/bioinformatic approaches (it is still important to do experimental structural biology and other types of computational biology), nor is it infallible, requiring logical scrutiny and experimental validation. But, at the same time, AF does change how things are done, even profoundly so, offering new insights (in a way, AF is now a research field of its own), and new ways of doing things faster or in a more high-throughput way, especially in the realms where it managed to infer some higher-level rules (standard protein modelling) and in other realms to the extent that queries resemble bits of training set (e.g. protein:nucleic acid interactions seem currently to robustly work only if there is homology to things already in the PDB, as far as I can tell from this report).
Similarly, the use of AI for maths seems to mainly work through ‘homology’ so far (finding existing but overlooked solutions in old literature, applying known methods to new problems, seasoned with some brute-force computation), although some higher-level ‘rules’ will probably be locally found in some types of problems, maybe already are, allowing somewhat more creative proofs to certain problems. With time tool limitations become apparent - and they probably will in the use of LLMs in maths - but the tool will have a profound impact, I am pretty sure of that (although the impact might also be partly negative, if it makes people lose their skills or critical acumen, or become demotivated, or focus only on things attackable with AI to the neglect of other, really new problems).
What always remains the most valuable is human attention: setting what is interesting, posing the question, critically evaluating/crosschecking, understanding and explaining to others the result, moving beyond limitations - and I think we will see that in maths too. When another revolution, cryoEM resolution one, took off in 2010s, what contributed a lot to progress alongside new tech was the old fashioned mol biol and biochem skills (sample preparation, optimisation) and the ability to interprete structural models, honed by the long tradition of experimental structural biology (the latter is useful for interpreting AF models as well). What fuels the AI revolution in maths seems to be the human ability to do and verify maths by hand.
Not to mention that currently most attempts to prove complex maths problems with AI yield incorrect results: you need ways to distinguish the rare correct ones, and understand their novelty (or unexpected triviality) etc. - something only an expert can do.
Lastly, if all this AI business, generally speaking, is really about human-collected and -created data, and whether methods and insights extracted from them extend to new cases – then it is clear that at some point one also needs new data, and every bit of new data is precious - in itself, and, secondarily, by adding to training sets. Doing things the slow way, manually, with reflection makes us notice and comment on things otherwise lost, hones our skills, gives time to think of other things in the background.
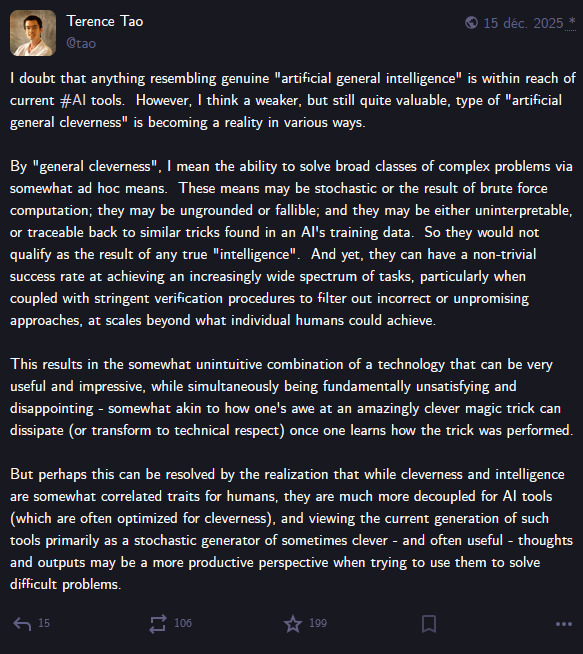
I leave you with a quote from Terrence Tao that summarises his intuition on the current AI in maths situation:
More filaments - with coiled-coils this time round (7/01/2026)
I enjoyed reading this study describing cryoEM structure of a NONO:SFPQ heterocomplex this morning, while it was snowing outside my window. How happy I was to read a paper in full (I have read only a handful of papers in full in the last weeks or even months, and some were to review). What a lovely thing to do: reading a paper…
Interestingly, the (endogenous, I think?) complex was natively purified from hamster ovary cells, as NONO has a native histag-like HHQHHHQQHH. Cryo-EM shows lovely two-stranded filaments composed of heterodimers connected via coiled-coils. For most other proteins we would need to knock in a tag to be able to do that.
We like filaments in our team, especially those of transcription factors. DBHS family of proteins to which NONO and SFPQ belong are not primarily TFs, but SFPQ does have a DNA-binding domain and has been suggested to activate and repress some genes.
We in our team believe that filaments are more common among TFs than it is currently thought. They might help with multivalent DNA binding or, conversely, sequester TFs away from DNA. Many other conceivable regulatory advantages.
A potential general lesson from this particular study: a lot of proteins (including many TFs) have coiled-coils, which are often cut out to facilitate purification and crystallisation. Coiled-coils are usually thought to mediate defined oligomerisation (dimerization, tetramerization), but I think quite often they mediate formation of much larger multimeric filaments or lattices. This is a lovely example.
Interestingly, the filaments seen in NONO:SFPQ Cryo-EM recapitulate contacts previous seen in crystals of these proteins – it does seem that, for filament-forming proteins, the crystallisation process often harnesses native multimerization properties. We like that, as we use X-ray cryst. a lot. But it is still vital to see the ‘solution’ structure with Cryo-EM – and in NONO:SFPQ case, it is only with EM that the complete superstructure and all the right contacts could be observed.
AI more generally (26/11/2025)
“Die Erfahrung entdeckt im Menschen, so tief sie eindringen mag, immer wieder nur Menschliches, in der Welt nur Weltliches, in Gott nur Göttliches. Und nur in Gott Göttliches, nur in der Welt Weltliches, nur im Menschen Menschliches.”
(“Experience discovers in human beings, however deeply it may penetrate, always and again only the human; in the world only the worldly; in God only the divine. And only in God the divine, only in the world the worldly, only in human beings the human”, translation by Barbara E. Galli, just with ‘human beings’ swapped for ‘man’ by me.)
Franz Rosenzweig, the famed German Jewish philosopher, wrote these words in his Das neue Denken, but I stumbled upon them in Hilary Putnam’s delightful little book Jewish Philosophy as a Guide to Life. I keep coming back to this quotation when reading yet another article about AI, for example, the short piece by a man from Google in the latest Nature (incidentally, it is interesting that this esteemed journal does not seem to realise the enormous conflict of interest when someone from a company investing billions in AI writes a quasi-philosophical text on AI).
What Rosenzweig means - I think - is that, in the end, the consensus human experience (with a possible controversy about God) is that these three categories - human, world, God - are different from one another. This one quotation cannot replace the whole history of philosophical and theological reflection that supports and sometimes qualifies it, but I like how forceful the wording is, and how elegant, particularly in the original German. And I think it is a good battle cry against the recent AI hype.
In recent months, I have been told, again and again, that if one somehow redefines the human being (as “intelligence” or “consciousness”, whatever they mean, or more bluntly as “something that, judging by outward appearances, might be taken for a human being”), then suddenly the difference between human beings and the non-human world disappears, and it turns out that I shall indeed find the human also in something non-human: in a computer. Well, I shall insist that I shall not find it there. A human being understands, knows, desires, strives, loves, regrets, and so on. His/her words really refer (a non-material, intentional relation) to external things (I am an Aristotelian in that respect). Machines do none of this, not even the most recent ones that are great at recreating apparent patterns. They generate appearances (more precisely: their makers generate appearances through them). Moreover, we understand very well, at a technical level, how machines do this - how they are trained and set up to do this in terms of their architecture. If Silicon Valley companies really believe they are creating a quasi-human, then - lest they admit they support slavery - they should stop forcing this quasi-human to perform tasks and to earn money for them. But this of course they will not do.
At the basis of our recognising other human beings as similar to us lies our own experience of life and a justified search for a similar experience, or at least some liminal form of that experience in the case of some animals, on the basis of certain external appearances. Yet whenever we attempt to infer some inner quality from external appearances that usually arise from that quality and are thus a reasonable proxy for it, the method works only so long as no one is deliberately trying to produce the external appearences without the inner quality. Otherwise the external appearences get uncoupled from the inner quality and become the target of their own (it’s like assessing scientists by citations, by the way). I guess what I mean is the same thing as what is more commonly known as Goodhart’s Law: “When a measure becomes a target, it ceases to be a good measure”. By the way, it is worth noting that it is seldom claimed that algorithms that only create images, like Midjourney, although they carry out operations of a similar kind to language models, are “conscious”. Such an illusion arises only in the case of language models because they deliberately imitate human utterances, use the first person singular, insert emotional words, print text gradually as if they were really writing and so on. Everything is aimed at producing a particular illusion, capitalising on innate human instincts to search for certain meaningful qualities (rather as in pareidolia).
These are some of my philosophical arguments. But on the practical, everyday level, what should one do? Using particular AI tools for particular purposes may be all right, might even be desirable; I use particular things that fall under AI (especially AlphaFold, other forms of machine learning) a lot in our scholarly work. I can see also how it can be useful in some tasks like coding or data analysis, although I have more reservations to that use. Yet most human uses of what is usually understood by AI seem to me a waste of precious human time and something that, especially on a broader, societal scale, has far more disadvantages than advantages. I would even risk advising the principle that, when in doubt, it is best to avoid ‘AI’ as much as possible, to use it as little as possible and to boycott it as much as possible. At least that is my modest private view.
A certain science-fiction writer (Frederik Pohl) is said to have remarked that a good sci-fi story is able to predict not the car but the traffic jam. Perhaps even better - I would add - if it were to predict people who spend the whole day in the car and then go to the gym to exercise, and at the same time the scale of changes in the natural environment caused by cars. I heard Bill Gates recently; he was saying how nice it will be when an artificial assistant remembers the birthdays of friends and rings up in advance to order a present or flowers. That is the car. And the traffic jam? That he forgot: florists overwhelmed by false telephone calls, recipients who see no value in these flowers.
The car: a machine producing grammatically correct and often factually correct texts. The traffic jam: people who are afraid to write and think on their own at work, and who spend evenings playing games so that the mind does not rust. Perhaps in the end it will be as it is with cars. They are useful for certain things, but on a larger scale, looking more broadly and more deeply, it is better to leave them parked and use as much as possible the two legs most of us have (if we can). On two legs one can do many things well and efficiently, especially when we make the effort. Now think what we could do with two hands and a head.
AI and science (08/11/2025)
I finally read the essay “Could AI Slow Science?” by Sayash Kapoor and Arvind Narayanan, computer scientists from Princeton and authors of the AI as a Normal Technology website (formerly AI Snake Oil). The main point of the essay is that even if AI might accelerate the productivity of individual academics, science as such is a complex system with emergent properties that are hard to predict, and here AI might actually slow things down. One could add to that that even the performance of a single individual is something complex, where, for example, a tool that gives a short-term gain (by speeding up certain seemingly mundane tasks) might lead to a long-term loss (by removing the opportunity to observe, think, and learn while performing those tasks), or something that accelerates performance in a particular area negatively affects other areas (e.g. the tool that makes writing certain unimportant texts faster might take from its user the fluency and self-confidence needed to write complex things the tool cannot really help with).
An interesting observation by the authors is that for complex processes such as scientific progress, accelerating certain aspects might have little positive effect – or even have negative effects – if these aspects are not rate-limiting to the process (“[it]’s like adding lanes to a highway when the slowdown is actually caused by a toll booth. It’s sure to make things worse”).
The first part of the essay is dedicated to arguing that – independent of AI – science has generally been slowing down in recent decades. I did not find this part that convincing. Science advances through breakthroughs – the authors say – not “minor improvements to known results”, and in recent decades we have had fewer breakthroughs, despite the huge increase in the number of published papers and scientific spending by governments. There might be something to this narrative, but I am not fully convinced. Sometimes science advances through large paradigm changes – what I guess we mean by “breakthroughs” or “disruptive science” – but in other cases, discovering more truth might actually be best achieved by going further in the same direction: testing, refining, applying, extending, and building upon existing paradigms. To some extent this would depend on whether a paradigm in question is on the right track and has high applicability potential. If it does, and arguably the 20th century’s scientific breakthroughs have led to some such fruitful paradigms in major disciplines from physics to biology, then it might not necessarily make sense to ask science to keep producing breakthroughs at any cost.
The authors sort of hint at these kinds of counterarguments, but do not buy them, one reason they cite being that we have been wrong so many times in the past in thinking we do not need fundamentally new ideas – and then, each time, new breakthroughs did suddenly arrive. There might be something to this logic, but I still think it does not necessarily make sense to assume science should always progress in the same way. This assumes certain things about the nature of reality and our way of discovering it that, I think, are not knowable a priori; rather – I would say – it is science and the way it develops that shows us what reality is like – how predictable or unpredictable, how multi-layered etc.
In any case, agreeing or not with their idea that science is slowing down is not really necessary to follow the rest of their argument. Even if it isn’t slowing down compared to the past, it could well be accelerated or improved – and it could well start slowing down in the future, which is what they say might happen when AI is more widely adopted by the scientific community. Their central thesis is that the slowdown of science is caused by overproduction, and this will be made worse by AI. I think we could transform this thesis into a simpler one: whether science is slowing down or not compared to the past, it is possible that simply maximising output – if that is what we are doing – might not be the best possible way of doing science, and AI might indeed make things worse.
The authors give interesting examples from coding and statistics. Coding practices, software documentation, and software maintenance are historically of poor quality in science, and so is the use of statistics. These things require competence from outside the given core discipline, and scientists might not have this competence at a high level, nor incentives to develop it, as they are just incentivised to keep publishing and keep discovering “significant effects”. This leads to subtle errors in practice (e.g. in the use of p-values) that might have large negative consequences. From this perspective – the authors say – AI might allow certain tasks – such as software writing – to be performed by scientists with even less competence in this particular task, making it even more difficult to spot and correct subtle errors. As evidence, the authors cite analyses of AI-enabled research performed to date, which seem to be largely flawed for various reasons (e.g. inappropriate baseline comparisons). The authors are, of course, aware that AI, if properly deployed, could potentially also help in spotting or removing errors. But on balance, it is not clear that it will help more than it will harm, at least if it continues developing in the current direction.
Another argument focuses on theory formulation vs simply being able to predict certain phenomena or arrive at answers in a black-box-like manner. The authors argue that formulating and testing theories advances understanding, while simply predicting, even if very accurately, or getting “answers”, might actually limit understanding (something apparently called a “prediction–explanation fallacy”).
Here, the authors cite an interesting essay by a Fields Medal laureate, mathematician William Thurston. In his essay, Thurston gives an example of a mathematical theorem that was proven using massive automated computation (before real AI arrived, but still with computers). Thanks to this approach, it was proven that this particular theorem is true. But it turned out that the reactions of other mathematicians to this proof were different from usual cases when theorems are proven in mathematics. There was less enthusiasm and reassurance. What people seemed to be lacking in this particular case was a human understanding of the proof (I guess this could mean the ability to yourself grasp the proof or - which might be equally important - to personally trust someone who says they grasp it). Thurston also gives another example: his former field of mathematics, which many of his colleagues “deserted” after he made several important proofs. People deserted it – speculates Thurston – because the fact that proofs were found (even if by a human) diminished the incentive to continue working in this domain, and perhaps also because – he admits – he described his proofs in a hard-to-understand way that discouraged newcomers from joining the field. In the end, the fact that this field was deserted resulted in what progress still remained to be made in this field being made more slowly.
These examples from mathematics might not be fully extrapolable to other domains – Kapoor and Narayanan admit – but some points seem valid to me. What we want is understanding, and this is made through diligent work on problems, not by just hearing the answers. And there is also the question of incentives and motivation, which might be eroded if what is often most exciting – answers – are produced by machines regardless of our efforts. Thus, even if AI does at some point get good at providing scientific answers (a big if, in my opinion), it does not mean it will speed up science – at least the kind of science in which humans still participate and which humans further develop and popularise. “If we use AI to bypass human understanding, or worse, retain only illusions of understanding,” the authors argue, “we might lose the ability to train new scientists, develop new theories and paradigms, synthesise and correct results, apply knowledge beyond science, or even generate new and interesting problems.”
This, I think, is an interesting point worth thinking about. Indeed, a lot of what we do in our research team, for example – all the team meetings, journal clubs, all the efforts put into teaching interns – all this has nothing to do with getting any short-term scientific “answers”. It is more about creating a community based on understanding, competence, and trust - a community that the society can then refer to when understanding in a certain discipline is needed.
And then, of course, there still remains the possibility that AI – understood as LLMs – will not even work that well at providing answers in the first place. This is not something that the authors discuss, but it is perhaps a digression worth making. I read recently about a few prominent mathematicians praising the usefulness of AI in their day-to-day work, so I do believe it might have use in mathematics, but so far the use has been mostly that of a very thorough search engine that finds, among all existing theories, lemmas, and proofs, those that a mathematician might be looking for but does not know exist. Quite likely, a similar use might be made more widely of AI in other scientific disciplines, to some extent facilitating the search of all existing knowledge. But more than that? I am not sure.
We have had relatively high-performing LLMs for some time now, and definitely for a few months. Trillions of dollars have been poured into them, almost everybody is using them. If they worked well in science, I think we would know. We would not have just a few anecdotal examples of people claiming AI arrived at complex conclusions they struggled to reach – but countless, well-documented, and validated cases of LLM arrival at complex conclusions. Anecdotal examples do not convince me that much. Even a wrong clock is right twice a day. A few months ago, I read about one researcher from Imperial who was wondering if Google Co-Scientist model had access to his mailbox – because, when fed with his papers, the model arrived at a similar new unpublished conclusion to the one the scientist in question had recently reached. It was an impressive, if isolated, case, and I wondered how many other scientists might have had similar experiences. Then some time (at least weeks) passed, and I heard one of the founders of OpenAI citing this particular case (and Co-scientist is not even from his company!) as proof that AI is good at science. So probably there was just a single case after all (or few cases).
All this applies to LLM-type AI. That more focused, more thought-through, and more task-specific machine learning–based automation trained on good experimental datasets might have a positive impact, I do not doubt – AlphaFold being a good example (but, precisely, with AlphaFold, unlike with LLMs, we knew very fast that it worked well). But when people speak of AI, they usually mean some kind of LLM-related “general” AI. And this I think might have little to offer after all.
But going back to the essay… What do Sayash Kapoor and Arvind Narayanan propose, then? What do they suggest? To avoid any sloppy, unprofessional use of AI, such as producing software without any prior competence, or poor literature summaries. Connected to that is the postulate to value and develop true human competence, including in areas such as software development. To change the system not to demand that scientists keep increasing superficial productivity metrics – as this will increase low-quality outputs that are easy to make with AI, leading to a deluge of low-quality science distracting us from the good one. And when developing AI tools for science, they suggest to consider making tools that help with real needs (e.g. looking for errors in code or data) or answer specific points, rather than producing global tools delivering flashy but suboptimal outcomes while killing human motivation and skills in the process. Finally, they advise a slow adoption of AI and monitoring of quality and emergent side effects – something we do not seem to be doing much.
Am I convinced by the essay? I was an easy audience, because I am generally sceptical of any LLM use, particularly in science, and quite attached to the idea that a human being – where properly (self)educated, hard-working, and kept to a high standard – can achieve a lot without too much machine assistance, and everybody is happier that way. Think of musicians in a good symphonic orchestra, for example. Am I convinced that AI will slow down science? Well, it could. Just being reminded that consequences of things in complexes systems are not easy to predict makes this essay worth reading.
Tim’s visit (22/07/205)
Recently, I hosted, in Orleans, my PhD supervisor, Tim Clausen, together with his family. It was a private visit, but we combined it with a visit to our institute Centre de Biophysique Moleculaire and our team, complete with informal and formal discussions with junior team members. I’ve spent 7 years with Tim (2011-2018), initially as a PhD student, and later as a postdoc. These have been formative years for me, in both academic and human terms, and Tim has been very important to me as a mentor. I’ve learned many specific, technical things from him - about protein purification, for example - but even more importantly, I’ve learned something fundamental about how to approach science. Tim has singular enthusiasm and daring when it comes to approaching scientific questions and formulating ideas. This enthusiasm, this daring - to the extent that I share them - I learned from him.
I’m a great believer in what Michael Polanyi calls the ‘tacit dimension’. ‘I shall reconsider human knowledge - wrote Polanyi - by starting from the fact that we can know more than we can tell’. Not everything that people teach us is transmitted through words or even easily put to words. There is this whole side to human knowledge - attitudes, practices, values, habits of mind and behaviour, etc. - that we learn not by listening or reading, but by observing, imitating, by immersing ourselves in a specific environment. I was lucky to learn from Tim and his team members, by being part of that team for several years. And now? I didn’t really believe it would happen, but there we are: now I’m myself a supervisor of younger colleagues and a co-leader of a team - well, at least officially, for really being a mentor or a ‘co-leader’ is, I guess, something one earns (or perhaps I should say: something that one occasionally succeeds in being, on rare, best mornings), not something that one just officially calls oneself. Be that as it may, thinking of the importance of the tacit dimension now makes me scared. Things that I now do and say, the atmosphere that I in part shape, the implied values, the unspoken rules - all this will shape my younger colleagues (while I am also shaped by my younger colleagues and learn from them). This is scary. But in the end, the best one can do is probably to keep trying one’s best - and to improve after mistakes and sins - trusting that it’s going to be all right. There’s only so much that we have control of. In any case, I was very happy that, being in this new position, one thing I could bring to my younger colleagues was to organise these short meetings and discussions with Tim, so that they can get to know him, talk to him. This is now part of their experience, their path.
But most importantly, it was a private, family visit, where our children could play with each other (without sharing a language in common!), where we could share meals, walks, discussions. That’s another ingredient of science that we often forget about - friendship.
Before a talk (20/06/2025)
Waiting in the park outside, watching people jogging, exercising. A young girl boxer is practicing with her coach, they are punching each other with boxing gloves. I’ve just passed a house with an inscription “Simone Weil philosophe a habité cette maison de 1929 a 1940” and tried taking photo of it with the zoom camera of my laptop, as I don’t have a smartphone, to send it to a friend who is very fond of her.
The second day of the annual SFBBM meeting in Paris won’t start until 8.30 am (it’s 7.20 am now), but I came from Orleans with the first train of the day just to make sure I’m not late as I’m the first one to give a talk today. The first day yesterday was very nice - a wide range of topics and good quality of presentations and posters, some familiar faces whom it’s a pleasure to see again and talk to. The meeting is held in the pharmacy department, one of the late 19th century’s “churches of science” meant to convey the spirit of positivism and scientism, complete with frescos and sculptures. The conference room itself is not in the best condition, the benches are a bit worn and the walls dusty, but that’s part of the charm of the meeting - that it is relatively modest, using public spaces used normally by student and therefore cheaper to organise. Behind the building there is a lovely small botanical garden.
I have a talk at 8.30 am and whenever I do, I sleep badly. I was woken up by my daughter at 3 am and didn’t fall asleep. Practiced my talk a bit, went to the train station on foot, took the 5-o’clock train, arrived just after 6 am, and then wandered around Paris for an hour murmuring the talk to myself. I made a new introduction - tried to make it a bit interactive - let’s see if it works out or not. I will speak about protein filaments made by ZBTB proteins and SIAH E3 ligases studied by us recently. I will ask people in the room to imagine they are proteins: if they can only use one hand to connect with each other, they can only make pairs, but if they can use both hands, depending on the angle between their two arms, they will make circles or chains. Et cetera. I will not ask people to actually do this but only imagine. It may be a complete fiasco, let’s see.
Will try to find some tea before it starts.
Springtime in the lab (14/04/2025)
Last week was beautifully sunny and warm. Spring has truly arrived, bringing new hope and joy. This week was supposed to be colder and rainier, and I guess it will be, but as I’m writing this, the spring sunrays are squeezing through holes in the roller blind. Hope and joy do not give in easily.
I’m alone for a few days - my family is at my parents-in-law - and my plan is to work a little bit more and finalise one manuscript. We have recently finalised another, smaller one. This year is a bit busy with things we need to finalise. But that’s good, too.
It’s my fourth year in France and in my permanent researcher job, so things don’t strike me as new all the time any more, and there is some sense of recurring yearly cycles, and it has a lot to do with what kind of interns we are having in the team. The spring, for example, is the time when the students from a vocational college (BTS) have just finished their internships, and the 1st year master students join for a two-month project, and the 2nd year master students are more settled into their work and have to start thinking of the writing. I’ve worked closely with one BTS student, one 1st year, and one 2nd year master student this year, and I’ve enjoyed it very much, even if it is sometimes a bit difficult to reconcile doing experiments with a student with other duties.
To continue with good things, our project about filament formation among transcription factors was selected for an HFSP Early Career award, meaning it will be funded. It was a second attempt, after failing to get the funding last year. Failing and having to reapply is never pleasant but it did the project some good - we have matured our ideas and made the project a bit more streamlined. Let’s see what we find.
Review on molecular principles of condensate formation (05/02/2025)
So there we are in Grenoble again, this time first on cryoEM data collection (today and tomorrow), and then on old good X-ray diffraction data collection (tomorrow and the day after).
Sitting and waiting for the data to be collected I have finally finished reading this review by Alex Holehouse and Simon Alberti. I liked it a lot: it is balanced, careful about overstatements or ambiguities, but also bold in making proposals as to how these things called condensates work, or how we might understand them, in an approximated but useful way, to work.
I liked the initial historical bit, bringing from oblivion old papers about ‘quinary protein structure’, and even more so the rest of the ‘Introduction’, where the authors defined the key concepts and distinctions. The biomolecular condensates emerge as molecular networks driven by the ability of some biomolecules to simultaneously form more than two (so at least three) homo- or heterotypic interactions (‘multivalency’). With monovalency you get dimers, with bivalency linear filaments, and with at least three interactions you get something that can start forming networks in 2D or 3D. ‘Multivalency’ is a key term here. Thus the controversial ‘phase separation’ just is partitioning due to network formation (so shouldn’t be that controversial after all, I guess, even to classically-inclided biochemists). The authors suggest it might be better to drop the common ‘liquid-liquid’ descriptor, as it is hard to formally demonstrate liquidity as opposed to ‘viscoelasticity’, i.e. some liquid and some solid features. I love how they give their due to all interaction types: from well-defined site-specific ones (between folded domains or a domain & a motif) to ‘chance’ interactions any exposed protein bits - in folded domain or IDRs - can form and which may be significant (particularly when things get locally concentrated), functionally relevant, and evolutionarily selected and maintained. To describe these latter interactions, they use the term ‘chemically specific’ (not sure I’m fully convinced), emphasising that they’re not encoded in specific folds or motifs but in bulk amino acid composition or permissive patterns.
As for the different types of site-specific interactions, they do mention multimerisation of folded domains, including filament formation, a topic of great interest to our team. And they cleverly comment that since the bivalency observed in filaments is not enough to make a network in 2D or 3D, filament-forming proteins will be expected to have additional binding properties as well.
One important fragment: “One common misconception is that IDRs are inherently predisposed to drive phase separation. This is no more the case than the idea that folded domains are inherently predisposed to mediate protein:protein interactions. In both cases, the key feature is the tendency of the proteins to undergo attractive intermolecular interactions, which is governed by the specific amino acid sequence”.
And another important quote: “While it is sometimes suggested that phase separation requires ‘promiscuous’ or ‘non-specific’ interactions, this is a misconception. While promiscuous multivalent interactions can drive phase separation, so too can highly specific multivalent interactions”. This last quote is very balanced, but the authors do allow hierarchy in condensate formation, with certain proteins acting as ‘scaffolds’ (perhaps more often through site-specific interactions with a dash homomultimerisation?), others as ‘clients’ that join - and necessarily also modify - the crowd, because what counts for final partitioning are all molecules, including ions etc. The focus on the review is on proteins, but they do discuss nucleic acids as well, and even poly(ADP-ribose), a nucleic acid-like polymer dear to my heart. The authors also caution against reading too much into in-vitro phase separation, as “almost any protein will - under an ‘appropriate’ set of solution conditions - be able to undergo self-assembly”. In vitro methods seem best adapted to studying robust site-specific interactions (which may contribute to condensate formation), but condensates themselves should be studied in cells, ideally quantitatively.
Lastly, it was just a pleasure to read. Alex and I did our undergrad together, especially in the first two years, sitting side by side in all tutorials, writing essays on the same topics. And now this - and all his phenomenal work on IDRs etc.! What a privilege to look back on those days!
Two condensate papers (17/01/2025)
The cell is not homogenous and, even disregarding the different specific organelles, it has areas of higher or lower concentration of various components, with many denser bodies or foci composed of proteins and other macromolecules. Thus, many cellular components are not evenly distributed but instead concentrate in distinct bodies/foci, despite these not being delimited by a membrane. We speak of the propensity to cluster or condense; we speak of condensates. And different cellular condensates vary in their composition - meaning there is specificity to the process.
I’m mostly interested in proteins, and more specifically, coming from structural biology, I tend to focus on folded protein domains and interactions in which at least one of the two partners is such a domain (the other can also be a folded domain, or a short peptidic motif that folds upon binding, or a non-protein molecule, e.g. a nucleic acid). Do such canonical interactions explain condensate formation? There are many examples where it looks like they do.
For example, we ourselves have worked (and continue to work) on protein domains that have an ability to homomultimerise in a chain-like manner, and this propensity is required for concentrating a particular class of proteins (e.g. transcription factors called ZBTB) into foci in cells.
I read two papers on condensate formation today, and they made me think of the various interactions that contribute to condensate formation, maturation, and dynamics. The first paper, by the Anthony Leung lab, is consistent with an important role for folded protein domains in condensate formation. The paper is about stress granules, cellular condensates that form under stress and are composed of various proteins, RNA, and a less common biopolymer called poly(ADP-ribose). It shows that a protein called PARP13 affects stress granule size and dynamics, and apparently does so thanks to its multivalent nature, with at least three different types of possible interactions, each mediated by a folded interface within PARP13. PARP13 thus homodimerises and can interact both with poly(ADP-ribose) – which seems key – and with RNA. As a consequence of its multivalency, PARP13 can act as a ‘crosslinker’ that provides vital connections within the noncovalent network that builds stress granules. These connections are not essential for the initial formation of stress granules - other interactions likely play a more fundamental role in maintaining the network - but they do influence the properties of granules, including their fusion and dynamics. This adds some nuance, by the way, to a simple distinction between the formation of a condensate or lack thereof.
The second paper I read today goes a bit against my instinctive focus on folded protein domains, but I don’t think it necessarily goes all the way to support the idea – sometimes entertained – that biomolecular condensation occurs primarily through some sort of nonspecific contacts between disordered regions. This paper is about the transcription factor FOXM1 and condensates it forms in cancer cells. It is true that the key FOXM1 region turns out to be not a folded domain but - by any standard of prediction - a disordered fragment. However, the core of the disordered region in question is not your typical disordered region: it is very highly conserved in sequence, can specifically self-associate with submicromolar affinity, and assembles fibre-like structures in negative-stain EM in vitro. I guess it is some sort of amyloid-like structure rich in beta-strands but more dynamic than conventional amyloids - if that is feasible? I tried AlphaFold and it does predict some sort of amyloid (but then AF almost always does when you ask it to multimerise a short peptide!). In any case, there is no canonical folded domain in sight, but the underlying self-association still likely is structurally defined, as the strict sequence conservation would be difficult to explain otherwise. By the way, the authors of the paper designed a peptide that mimics the key region and which, by interacting with FOXM1, interferes with condensate formation, with beneficial effects in cells and in vivo.
Thus, with these two papers, we are still in the realm of either folded domains or defined, conserved self-interacting motifs. I guess you could have condensates driven by less well-defined (‘fuzzy’) interactions - but you would still need these interactions to be somehow specific, if they really drive specific condensates. And in the protein world, specificity usually, sooner or later, boils down to specificity for particular amino-acid residues (or classes of amino-acid residues) in particular positions, even if these do not form a contiguous motif but are distributed on a surface of a folded domain or as a specific pattern or ‘grammar’ within a disordered region. I do think there might eventually also be a role for less specific, very transient interactions mediated by disordered regions not conserved in sequence - but intuitively I would see such interactions as auxiliary, happening between partners that are already brought into proximity through other means.
I should finish by saying that these are just some loose musings prompted by reading and by extrapolating a structural biologist’s thinking to cell biol… I know there are actual experts working on these things. A friend, Alex Holehouse, has just co-authored a review that seems to cover these topics and which I will try to read next.
Nonscientific reading 2024 (06/01/2025)
In 2024, I kept, for the first time, I think, a clear record of all booked I read, in a chronological order. For many years now, I have a diary, where I write quite a lot every week and I also write on books, but now I added a list where I put down the titles I’ve read at the back. It’s hard to compare with previous years, but it seems like 2024 was a decent reading year for me — at least compared to other years since starting a family. I don’t have much time to read nonscientific things these days, and most of it happens on the tram during my commute back from work or a bit before bed. I try to keep notes on my reading in my diary, which takes some extra time. For this list, I’ve included not only things I read by myself but also a few longer children’s books I’ve read with my daughters (two by Kästner and one by a forgotten Polish author Buyno-Arctowa; we’re currently reading another Kästner). The books are a mix of Polish and English (some of them translated into one of these two languages from another). There wasn’t a specific theme or method to my choices — reading has become a “sphere of freedom” for me, where I try to just follow my intuition or chance. That said, I also try to read books that are gifted or recommended to me.
One book that stood out for me this year was a biography of the film director Éric Rohmer. It allowed me to dive into an art form I know little about and explore one creator’s life and work. My wife and I also watched some of his films available at our local library. I’ve written about it in one of the posts below. I’m glad I tried Trollope this year, an author I hadn’t read before; I’d like to read some more. I also enjoyed the first volume of Stefan Swieżawski’s memoirs (he was a historian of philosophy from Poland). There are two more volumes to go, which I hope to finish in 2025. Some books were short (which can be motivating when keeping a reading list!), but two were longer — the one on Rohmer and the one on Péguy.
Does nonscientific reading have any impact on how we approach science? I like to think it does. At the very least, it helps with writing. But I believe it can do more: it can shape the way we think, structure arguments, and imagine models and metaphors. It can also teach us to observe more keenly, nurture curiosity, and even offer lessons in morality — and I do think morality is one of the key things in science. One of the books I read this year was an essay by Christopher Dawson on the beginnings of the Oxford Movement. For those unfamiliar (it is a bit of a niche topic, I guess), the Oxford Movement was an unusual episode in the history of Anglican theology, marked by going back on some of the precepts of Reformation. Dawson highlights that the movement’s courageous pioneers (well, one of them in any case) were/was inspired by the medieval ideal of chivalry — understood as a selfless and brave dedication to a chosen goal and moral integrity, even in the face of difficulty and criticism. This ideal could be inspiring for scientists as well, couldn’t it? I think there are interesting lessons for scientists in Rohmer and Péguy, and Naipaul is a good teacher of concise style. And Trollope’s portrayal of a career in politics — where ambition meets morality — has many parallels to the world of science.
Nonscientific books read in 2024:
Éric Rohmer: A Biography, Antoine de Baecque & Noël Herpe (English, trans. from French)
Tajemnica Frontenaków, François Mauriac (Polish, trans. from French)
Anatolin, Hans-Ulrich Treichel (Polish, trans. from German)
Uległość, Michel Houellebecq (Polish, trans. from French)
Emil i detektywi, Erich Kästner (Polish, trans. from German)
Miguel Street, V. S. Naipaul (English)
Fantomy, Maria Kuncewiczowa (Polish)
The Spirit of the Oxford Movement, Christopher Dawson (English)
Carnal Spirit: The Revolutions of Charles Péguy, Matthew W. Maguire (English)
The first three volumes of Tintin (French)
Mania czy Ania, Erich Kästner (Polish, trans. from German)
Wielki przełom: 1907–1945, Stefan Swieżawski (Polish)
Kocia mama, Maria Buyno-Arctowa (Polish)
Nie mówię żegnaj, Han Kang (Polish, trans. from Korean)
The Autobiography, Anthony Trollope (English)
Can You Forgive Her?, Anthony Trollope (English)
Seeing before knowing how to do (11/12/2024)
I have stumbled upon this graph, by a modern realistic painter Marc Dalessio, from his website. It is about a situation where your ability to paint better has not yet developed, but you can already see more than before and, therefore, are more critical of your current work. According to him, such moments of increased self-criticism generally precede moments of improvement - for him as a painter, at any rate. If seeing differently is actually required to paint differently - but is not by itself sufficient and requires the hand to learn to follow the eye, so to speak - this seems to make sense. I thought that it might apply to scientists as well, offering some comfort to those who are plagued by frequent bouts of self-criticism, as many scientists probably are. Being self-critical assumes we already sense something but do not yet know how to address it - perhaps we sense our writing is not good enough, or our way of asking questions, or a simplistic model we assume, or a simplistic methodology we use. It is comforting to think that such moments might generally precede improved ability to match the increased expectations. The problem is - it never stops…
November update (23/11/2024)
The last time I wrote anything here was just before the EMBO ubiquitin conference held in Cavtat, Croatia. The conference marked an important moment in my life, as it was the first larger conference at which I presented the work done by my colleagues and myself since I started my permanent position in France. Also, and perhaps more importantly, it was the first confernece to which several of us - members of my current research group - went together. Going together as a team to a conference is always special, and when you are partially responsible for the scientific direction of the team, it is extra stressful but also more of an adventure. We shared two little apartments we rented, and the walk to and from the conference venue. We sat next to each other in most talks, then discussed what we heard. The place was beautiful, the atmosphere - welcoming and exciting. I saw some people I’d seen before, others that I’d only heard about, or read their name on publications - so that you can put a face, so to speak, to the name and to the science that you’ve known. And I’ve met some old friends, particularly from the group where I did my PhD. I avoid travelling not to abandon my family, but I do always find it surprisingly enriching whenever I do go somewhere. There have been a few other travels that I have done since then, mostly small and with a maximum one-night stay. First - this year’s edition of our Biotechnocentre meeting - the regular event about which I’ve written here already once, because I’m particularly fond of its mixed scientific-social format. And then, just last weeks, I went to two small French meetings, one in Strasbourg and one in Paris Saclay. It was snowing during the second meeting, the buildings on the other side of the large window of the lecture hall growing progressively white as the talks went on, the snow giving the occassion a somewhat festive feeling.
I have my habilitation defence next week - it makes me very scared, but, in some way, I’m looking forward to it, particularly to the social aspect of it - meeting people, talking, hopefully celebrating a little bit. I’m told it’s a formality, but I’d like to use it as an occassion to mark our move as a family to France and the first three years of living and working here.
I saw a little video the other day of an actor, I think it was Alan Rickman, the guy who played Snape, giving advice about how to be a better actor. What he said, or in any case what I remember from it, is that to get better at acting, you shouldn’t limit yourself to studying acting. You should go to museums and read books, and do other things that make you grow as a person. And then, out of that, the acting might get better as well. I found it wise, although there should probably be more emphasis not just on intellectual, but also on moral development. And, of course, I do think that this sort of advice applies to science as well. If we want our science to get better, what we need, I think; is not just studying science, but studying, reading, living, doing other worthwhile things that make us more of a person. And, out of that, the science might just get a little better as well.
Binding regions in ignored flexible protein tails: a case of XRCC4 and XLF (26/09/2024)
Just finished reading this paper, which I think is a very interesting case study in protein biochemistry. The authors looked at, previously largely ignored, flexible tails of two DNA-repair proteins, XRCC4 and XLF, identifying in these tails various specific binding regions. These regions, mapped at aa resolution, were shown to mediate either homotypic interactions, or heterotypic interactions (with DNA or with specific proteins that belong to the same DNA-repair complex). These interactions are transient/weak but could become significant, when the partners are brought together through more canonical structural interactions. In the end, this increases the number of possible binding events between the partners of this DNA-repair complex. As these events can happen within as well as between individual copies of the complex, they can facilitate clustering of multiple such complex copies together. This kind of protein condensation is in agreement with classical biochemistry: specific binding events, measurable affinities, avidity and cooperativity effects. As the core protein:protein interactions (those that are relying on highly conserved interfaces and motifs) are getting more predictable with computational approaches, I have a feeling we will have to focus more and more on such ancillary contacts, which might be more plastic in evolution and less predictable. NMR seems perfectly suited for this (and I’m saying this as someone who has never used NMR) - it would be nice to use some NMR of that kind for our projects! Are these ancillary contacts functionally important? The authors show that when you get rid of them, you do get a slight if measurable effect on DNA break repair in a cellular assay, and the effect is much greater if you additionally destabilise the complex. This points to these weaker interactions as increasing robustness of the system, which I can easily imagine is important, if not in an isolated cell, then at some point in the development of complex organisms, leading to the relative conservation of the identified interaction regions. Congratulations to the authors of this very interesting work, which was a pleasure to read as well.
Protein filamentation (23/07/2024)
It’s been already a week since our paper on filament formation by the BTB domains of some ZBTB-family proteins is out, but I’ve only just come back from holidays. For more details on our paper, see the summary that I’ve just posted in the “Publication summaries” section of my website. Here follow more general comments about protein filamentation.
When we hear of ‘filaments’, we typically think first of cytoskeletal filaments, which are abundant, long, and are (dis)assembled in a regulated manner. But many other proteins with a tendency to homomultimerise in one dimension are known. This is often called ‘polymerisation’. I prefer to speak of ‘filament formation’ or ‘filamentation’, because polymerisation implies a particular size range (poly as opposed to oligo), but what is really distinct about these assemblies is their shape and topology. ‘Dynamic filamentation’ refers to a particular subtype where filaments (usually of limited size) form and dissociate spontanously without additional factors. Such are the filaments formed by BTB domains that we characterised, such are the filaments previously reported for domains such as SAM or DIX.
Filament formation is ultimately a question of topology/geometry: if the homotypic interfaces are located on a protein in such a way that you can always add one more subunit without potential clashes, you have a potential filament. But whether such a filament will exceed a few subunits depends on affinity & local concentration. There are many geometries that could give rise to filaments. The best studied is ‘head-to-tail’ geometry: the front of a protein domain interacts with the back of another copy of the same domain. See this lovely review by Mariann Bienz, one of the pioneers of the field, on protein domains that make filaments with the head-to-tail topology. In the case of the BTB domains that we characterised, the topology is different: they first make head-to-head dimers (which, BTW, is the most easily evolving and abundant type of homooligomerisation - see my post about protein homodimerisation at the bottom of this page for details), and then the dimers connect with each other through tail-to-tail contacts.
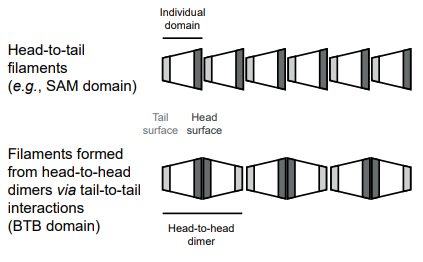
There are other topologies that give rise to filaments too. All this is important for their evolution. And also for prediction of filamentation with AlphaFold. AF2 or AF3 can work (doesn’t always do, but can), but has to be done in a way consistent with the expected topology. Head-to-tail polymers can be predicted by modelling a dimer: a protein might be a filament if you get a head-to-tail dimer with an angle allowing extrapolation to a filament rather than a ring. This is the clever approach taken by Emmanuel Levy’s lab and colleagues in the recent AlphaFold2 prediction-based study. For filaments built of dimers you need to model at least a tetramer. And so on. We comment on this in the discussion of our paper. Moreover, I think that in addition to 1-dimensional filaments, there might also be systems where proteins homomultimerise in 2 or 3 dimensions.
Is filamentation more common than we know? I think so. 1) They emerge easily in evolution (see this fascinating paper, again from Levy and colleagues). 2) They could explain some instances of foci in cells. 3) They might have been overlooked, because filament formation often causes insolubility on overexpression. This classic paper is a good example of the last point. The domain was insoluble, they fused a fluoresc. protein and did mutagenesis, found a solubile mutant. The mutant crystallised as a filament with the mutation at the interface, weakening the filament & making it soluble. Which, by the way, is also a good illustration of why crystallisation is a useful technique for filaments: crystal contacts might recapitulate real homotypic contacts, even if these are weakened by a solubilising mutation. But one has to be careful. Some crystal filaments might be a crystallisation-induced transformation of a ring-like multimer (see e.g. P65 or P61-symmetric crystal forms of ATPase domains). And being careful is generally a good idea when dealing with potential filaments. One has to think of artefacts of overexpression, etc.
A key technique for studying filaments is, of course, electron microscopy. As filaments are generally helical (you have to be helical or linear; otherwise, your multimer will circularise sooner or later), helical reconstruction of EM data comes in handy. SAXS and AFM are also useful in our hands.
One might add reason no 4 to why filaments could be more common than we think: because they are potentially functionally interesting. They concentrate a protein (in a specific manner!); they can give rise to avidity & cooperativity effects; they can create large structures with emergent physical properties.
Some general things we have noticed when working on filament-forming proteins (ZBTBs and others): 1) problems with solubility & aggregation, 2) predictability with AlphaFold2 or 3, but not always, 3) submicromolar or around micromolar affinity between protomers, 4) be careful with fusing not monomeric GFP as it amplifies foci (on the other hand, this might sometimes be useful for screening purposes).
CBM retreat, 16th-17th May 2024 (04/06/2024)
We had a two-day retreat of our institute, the Centre de Biophysique Moleculaire, held in a resort about 45 minutes away from Orléans, in the Sologne. Have you read the book Le Grand Meaulnes (translated several times into English, each time under a different title). Well, Le Grand Meaulnes is set in the Sologne, a region of forests and ponds south of Orléans. It was the first retreat since I joined (which was in 2021) and, in fact, the first, I’m told, that has been organised by the CBM in 9 years. We had scientific presentations by all the departments, some administrative stuff, long meals, a party. All in all, it was a lovely, joyful occassion. And the nicest thing, perhaps: a little surprise event for our long-time maintenance man, Justo, celebrating his many years at the CNRS with some videos and speeches sent by former colleagues. He, and two other long-time maintenance/administrative employees, Patricia and Martial, were awarded one of the medals of the CNRS for their contribution to the life of the institute over many years. A very moving occassion.
Alain Nicolas symposium, 11th March 2024 (09/04/2024)
I’ve been meaning to write something about a one-day symposium that I attended in Paris the other day, which celebrated the long career of Alain Nicolas, a French geneticist (who is still active and has recently moved from Institut Curie - where the event was held - to Niece). Now when I finally have a moment to write a few lines, unofrtunately, I don’t have my notebook with me, in which I jotted down some of my impressions. Anyway, I was quite impressed by this occassion and very happy to have decided to register for it in time, despite not knowing Alain Nicolas personally and not being from the field (genetics of recombination). It was a family event, in the sense of a scientific family: the speakers were recruited from among Nicolas’ coworkers (including his postdoc supervisor, the Noble-prizewinner Jack Szostak, who joined via zoom; several other American collaborators were there in person though), trainees, and colleagues. I could sense that we are celebrating someone special, a dynamic and inquisitive scientist, a good organiser, a faithful collaborator, one of the leaders of his field. Nicolas is a heir to a double heritage: the French one, that of the geneticist Jean-Luc Rossignol (Nicolas organised an event to commemorate him when he died in 2016), and the American one - that of Jack Szostak and a number of other figures whom Nicolas met during his postdoctoral stay in the Boston area. Someone mentioned that Rossignol, from whose lab in Orsay (near Paris) Nicolas originates, was, at the time, universally famous among geneticists, at least those working on recombination, and once he came for the first time to a conference in Cold Spring Harbor, everyone was intrigued, knowing him previously only from publications. He worked on the model fungus Ascobolus immersus, but Alain Nicolas moved to yeast and then I think also human cells in his later career. What a pity that Rossignol remains almost forgotten outside his direct circle now - it’s difficult to find anything about him on the internet.
In a way, this day was an exposure to the kind of science that is going away: science forged during conferences and through strong bonds between relatively small groups of scientists (some of them with imposing personalities) working on a given problem. Sure, conference still exists, but all the fields are much larger, the connections weaker. There were probably also negative sides to the way science used to be done compared to now, but this was the day to celebrate the good ones and learn from them. I took a lot of notes - scientific and also just ‘sociological’, if you like - and came back to Orleans very happy.
An additional reason for enjoying the symposium was that it was held in (or in any case next to) Institut de biologie physico-chimique (IBPC), an institution about which I have read a lot in the past. Founded in 1930 by Jean Perrin (a physicist and Nobel laureate) with the financial support of baron Edmond de Rotschild, it served a bit later as a model for the CNRS, the huge institution in which I happen to work, but is also interesting in itself, or rather through some of the personalities working there. I’m thinking especially of Louis Rapkine, an enigmatic, but apparently very amiable, researcher who sadly died young in 1948, and is known not so much for his research but for helping the exiled Jewish scientists, for his passion for culture (literature, music), left-wing, humanistic, but also somehow spiritual leanings. I remember reading a sort of obituary or memoir about him written by Leon Edel, the biographer of Henry James - I have to find it again. Outside the IBPC/university campus where the symposium was held, in Rue Pierre et Marie Curie, there were posters/panels about the history of IBPC, which I have now found online.
Filmmakers and scientists (29/02/2024)
I took a week of holidays to spend it with my family. During that time, I managed to read a little bit and finished a book I had started a long time ago, the biography of the French film director Éric Rohmer (1920-2010) by Antoine de Baecque and Noël Herpe. I don’t know how long it had taken me to read it, but probably several months. Mainly for my lack of time and the sheer length of the book (around 550 large pages), not for it being boring. Not at all! On the contrary, it was really enjoyable and made a nice break from science. I read it in the English translation published by Columbia Uni Press. I don’t know about you, but for me pastime reading is the one enclave of freedom, separate from work and any other duties. Having said that, I would say that filmmaking, at least of the type that Rohmer practised, is not that far from science; and neither is, in fact, any good art - a thought, perhaps naïve, to which I will return at the end of this blog.

My reading Rohmer’s biography has gone along with watching a few of his films (found in our local library) every now and then over the last several months. We have a few more that we would like to watch. I guess my main motivation has been that, now living in France for the first time, I feel like I should learn a bit more about the French culture, including the 20th century, of which my knowledge is rather patchy (I like some writers and some philosophers, but I don’t know much about more popular culture). Films from the French new wave felt like a place to start, and of the directors I have read about, Rohmer felt like a kind of cinema I would like. And I did like his films, a lot. My wife, for the record, much less, and it’s a testament to her love that she watched them with me anyway. She found the films a bit imperfect, amateurish. For my part, I like the way de Baecque and Herpe put it in Rohmer’s biography: “Rohmer preferred the actor’s irregularities that gave his story the imperfection of life”. “The imperfection of life” - that’s it!
It’s hard to describe Rohmer’s films to those who haven’t watched them. People talk a lot in his films, and live through small dilemmas of everyday life, often connected to relationships. Rohmer is someone who likes people, is curious about people, about how they behave and react, and he just shows that, in a fairly simple but somehow captivating way. My favourite of his films so far has been The Green Ray (released one year before my birth) - a purposefully banal story that nonetheless makes you think (about the interplay of chance and purpose, for example) - and smile.
A strange question perhaps, but one that motivated me to write this blog: is there something that an aspiring scientist like myself, a young scientist who might fail any moment but otherwise enjoys science, can learn from Éric Rohmer in terms of how to do science? I think there is, based on an analogy between Rohmer’s filmmaker’s practice and career and that of a scientist’s. Films, like science, are made in teams, for example, and Rohmer seems to have been someone who was good at bringing people together through rituals (5 pm tea breaks), through discussion on any topic whatsoever, through friendship. Also, one needs to get funding to make films, just like one does to do science, and getting funding is fuelled by recognition. Most films that Rohmer made were done quite cheaply. Being economical and careful gave him freedom, because it was easier to collect modest amounts of money, and the pressure for returns was smaller. In terms of recognition, Rohmer generally kept doing what he thought was good cinema and didn’t seem to try to please others. He was sometimes ridiculed and often misunderstood. Perhaps he didn’t take feedback sufficiently into account, but there was also virtue in his faithfulness to his own vision and values. He did make some films that have been judged as failures. But also many that have been acclaimed by critics and loved by cinema-goers. It’s a lesson in not caring about recognition but just trying to do good art. Or science, for that matter. If needed with modest means, but strong values and a sense of purpose.
When reading the biography and watching Rohmer’s films, I have been struck by these similarities between a filmmaker’s and a scientist’s fate. But I also kept thinking about a more general unity of science and art. Isn’t filmmaking and perhaps most other artistic activities about capturing, naming, and/or showing something of reality that has not been captured, named, shown before? That seems to me to be on the same continuum as science. Moreover, quite often – as in the case of Rohmer’s historical films, for example - it takes a lot of actual background research (of historical but also technical nature) to do art, in addition to artistic sensitivity and skill. Now, I like a lot a philosophical approach called virtue epistemology, which teaches that gaining knowledge and discovering truth requires certain virtues and skills, some of more moral kind, like selflessness or humility, other more “technical”, like perceptiveness.

In a volume of essays from this genre entitled “Virtue Epistemology Naturalized: Bridges Between Virtue Epistemology and Philosophy of Science” (Springer 2014), there is one by Shannon Vallor, who speaks of “perceptual responsiveness” as the basis of good scientific observation. Vallor, a philosopher from Edinburgh, gives the famous 20th century geneticist, Barbara McClintock, as an example (another is a pioneer microscopist from the 17th century, Robert Hooke). This is the description of scientific practice that Barbara McClintock gives: “You are so absorbed that even small things get big… Nothing else matters. You’re noticing more and more things that most people couldn’t see because they didn’t intently go over each part, slowly but with great intensity. It’s the intensity of your absorption.” And elsewhere, complaining about the attitude of some scientists: “I feel that so much of the work is done because one wants to impose an answer… They have the answer ready, and they want the material to tell them”. Anything else it tells them, “they don’t really recognise as there… If you’d only just let the material tell you”. These are all words by the geneticist, Barbara McClintock, who, through this kind of openness to reality and resistence to preconceived ideas (but also through her technical skill, familiarity with her specific material, i.e. maize, her mathematical analysis, her sheer hard work) could provide a description of the biological process of gene transposition. But these words could, just as well, have been written by a writer. Or a filmmaker. Indeed, to me, this kind of openness to reality is what brings scientists and artists together, definitely artists like Rohmer. Of course, the aspects of reality that artists are usually interested in are different from those that interest most biologists. Artists, writers, filmmakers are more interested in human relationships, in feelings, in our experience of the world and ourselves, in things like chance, fate, and providence, in what it means “to be rather than not to be”, in the moral complexity of human actions, etc. Psychological, metaphysical, moral questions. But the basic tasks of seeing, naming, showing the multi-layered and multi-dimensional reality in which we live are not that different. “It’s not myself I see in my films, it’s the world I’ve filmed”, said Éric Rohmer.
Inhibition of SUMO1 by its own N terminus (26/02/2024)
We started a journal club a few months ago. One meeting a month. In the 3rd session, we discussed the recent pre-print about the role of the flexible N-terminal tail of SUMO1 in regulating SUMO:SIM interactions. It’s a very nice combination of biochemistry, NMR, and simulations (and a little bit of in vivo). The prepring is here. There are two main molecular differences between ubiquitin and its homologue SUMO: 1) SUMO behaves like a beta-sheet that misses a beta-strand and it can interact with SUMO-interacting motifs (SIMs) that bind in a beta-strand conformation; 2) SUMO has a flexible N-term tail.
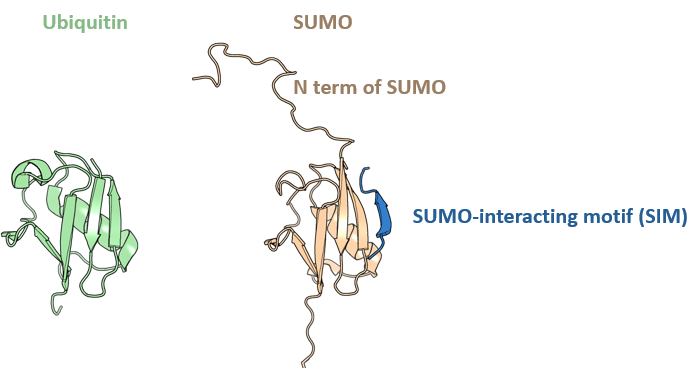
I should add that in humans there are 3 main paralogues of SUMO, with SUMO1 being a bit different from SUMO2/SUMO3. All bind to SIMs. The basic idea of this study is that the N-terminal tail of SUMO1 (but not SUMO2/3) inhibits the binding of SIMs to SUMO, providing regulation.

The inhibition seems to be due to a transient, ionic intramolecular interaction between the N-terminal tail and the area around the SIM-binding grove. This is nicely suggested through NMR, mutagenesis, and molecular dynamics. This is conserved in the sole S. cerevisiae SUMO Smt3 and in the C. elegans SUMO SMO-1. It doesn’t extend to the human SUMO2, but the authors show that, probably, if the SUMO2’s tail gets phosphorylated, it will get the right electrostatics to start inhibiting SIM interactions. It was my turn to present this time. Since one of the last authors of the paper is Frauke Melchior, one of the discoverers of SUMO, I made a short historical intro including this nice interview. I recommend the paper & congratulate the authors (and thank our journal club attendees).
Being a young scientist (26/02/2024)
I’ve read two interesting advice pieces written by established scientists, How to be a good Principal Investigator by Lawrence Banks and Things you should learn in graduate school by Ramanujan S. Hegde. I’ve been once in a (purely scientific) talk by Hegde and was very impressed; I’ve never heard or seen Banks, but I know of his work in virology. They belong to different generations and one can sense it a bit in the two texts, but both articles are interesting and useful. I’ll write a few things down here, not to forget them.
Banks’ article is meant specifically for group leaders. He divides his advice into three parts, tailoring it to new PIs, mid-career PIs, and late-career PIs. The advice is very different depending on the career stage, but there are some common threads. Overall, there is an emphasis on providing motivation to the team, on choosing the team well (so that older students are role-models to the younger one), and on leading by example (which, for new and ideally also late-career PIs, includes keeping working at the bench). Team building through regular coffee breaks or after-work get-togethers comes through as more important than regular group meetings, which Banks doen’t particularly value. It’s more fruitful, he says, to create an atmosphere in which people are eager to share and discuss with others their results as these are “hot of the press”. “Foster an environment - Banks advises - where lab members are comfortable coming back from developing a western blot and sharing their results: ‘wow guys, look at this! - this is the second time I’ve done this and the result is exactly the same - this is completely unexpected and something really new - what do you think?’”. One way to ensure such an environment is not to let the lab grow too big, beyond what is managable; and to always have time for your people, even when you are a “late-career PI” busy with departmental duties etc. Banks emphasises a few times that being a successful scientist involves working very hard, also at weekends, and that students should ideally come to understand that by themselves. This was where I wasn’t sure if I agree. Working hard, yes, but perhaps within reasonable limits. I don’t know if I’m a successful scientist, but I’ve almost never worked at weekends in my life, and I’m grateful to my mentors for letting me not do it. But I understand that Banks’ experience is probably different.
Now Hegde’s article, which is technically meant for PhD students, but I think it’s a very valuable reading to any aspiring scientist. His main message is that more important than mastering any particular technique is acquiring a “scientific instinct”: a skill that includes finding problems that are important, interesting, and doable within your constraints; then dividing the problems into short-term projects; then knowing how to learn the techniques that one needs to execute the project (the answer is: learn enough about the technique in question to be able to customise it to your specific question); and, finally, presenting the results in simple, logical, compelling prose. Because scientists, to stay productive and successful, are obliged to often switch projects or even fields and adapt to constantly changing techniques, Hegde’s emphasis is on what is enduring and transferable. A PhD student usually begins with a project allocated to them by an advisor but should ideally move to a position when she or he is able to propose important and feasible project by her/himself, and not only in your direct area of research but also more broadly. Hegde argues that the “scientific instinct” is not some irrational intuition but the ability to act in a reasoned and logical manner in the world that is full of uncertainties and where information is always incomplete. Thus what seems instinctive in some exceptional scientists is probably highly logical and reasoned, it’s just that the rationale remains unarticulated. And you can learn from such exceptional scientists, perhaps by encouraging them to articulate how they reached that particular decision. Generally, we should - Hegde says - learn from others, not just our direct supervisors, but any people in whom we notice qualities that we would like to develop ourselves. In addition to mentors, we can also learn by examining how some specific decisions have been reached in the past - those leading to failures and success. One interesting point that Hegde makes is that successful scientists often are skilled in making inferences from analogous problems in other areas - which means you have to read widely also outside your domain. And finally: when writing an article, you must put yourselves in the shoes of someone who doesn’t know your reasearch, and you must adapt your narrative (composed of simple, logically connected paragraphs, each containing a single main point) in such a way as to logically argue how your results support a given hypothesis. Since it has to be a consistent narrative, Hegde doesn’t see a point in starting with describing results or making figures first; rather, he says, write in the same order as the one in which the text is read, from the title to the discussion, adding and adapting figures to the linear narrative.
I think these two texts, very different and aimed at different audiences, and written by authors from different generations and with different personal experience are actually quite complementary, one emphasising mainly the human and social aspects of a good environment, another - the individual skills one needs to develop. I’ve found both texts really illuminating. Of course, in the end, science is not only about science, and not only about success (indeed, being a good scientist, or at least a decent human being who also does science, often means one needs to fail). Science must be reconciled with other important areas and other domains of human and social life, particularly with the world of values, justice, kindness, morals.
Modelling oligomerisation with AlphaFold (19/02/2024)
Paper from Emmanuel D. Levy and his group are always very interesting. This study, now published in its final version, is about modelling protein oligomers in a high-throughput manner using AlphaFold2. It’s worth a read, not only because of the excel file with predicted oligomerisation symmetries for most human proteins and those from three other species (E. coli, P. furiosus, S. cerevisiae). The approach the authors took to predict oligomeric states of proteins with AlphaFold2 with reasonable computational resources was to first model dimers, analyse the confidence scores, and - if those were sufficiently high - analyse symmetry of the dimers. Dimers with head-to-head (C2) symmetry were identified as potentially just dimers, while dimers with head-to-tail symmetry were identify as potentially ring- or filament-forming. By extrapolating the head-to-tail dimeric models, possible numbers of protomers in a ring were proposed (trimer, hexamer, decamer, etc.), or - if the oligomer didn’t close into a ring - a protein was suggested to form a filament. Interestingly, a standard version of AF2 - trained on single protein chains - was used instead of the ‘multimer’ one, as the gain with the latter was apparently limited and to avoid the situation where the training PDB set contains quarternary structures that the study predicts. Like always in the Levy lab papers there are many interesting general observations, like e.g. that the number of possible tertiary folds seems much more limited than the number of possible quaternary arrangements - AF2 identifies many new quaternary but few new tertiary structures.
One should note that not all possible oligomeric architectures are covered by the adopted approach: higher-order arrangements built of symmetric C2 dimers would not be predicted, for example.
All in all, it’s a very interesting paper with a lot of new information.
PARP1 condensates (12/02/2024)
PARP1 is the DNA-damage sensor that binds to DNA breaks and synthesises protein modification made from long poly(ADP-ribose) (PAR) chains. It is also my friend since my second postdoc, when I worked on PARP1 and its interactor HPF1 - a very interesting protein, by the way, which forms a composite active site together with PARP1 and changes the way it makes PAR chains.
I try to follow the literature on PARP1, especially as we are trying to do some work related to it in my current group as well. Among the different papers I have read on PARP1 lately, the recent one in Cell from Simon Alberti’s group has been the most interesting. Here is the link. Really, a very interesting paper with elegant biochemistry. Upon binding to exposed dsDNA ends mimicking a double-stranded DNA break, PARP1 can multimerise at relatively low concentration into large isotropic complexes that manifest as condensates observable under microscopes. The PARP1 multimers formed on dsDNA binding are mediated via specific interactions between structured domains from different PARP1 molecules. Some of these domain:domain interactions had been proposed before, occurring in cis within the same PARP1 molecule, but it looks like they could occur in trans between different PARP1 molecules, thus non-covalently cross-linking multiple PARP1s into larger objects, beginning with a dimer. There might be further unknown interfaces needed for this process (and perhaps some are hidden as crystal contacts in existing crystal structures?). PARP1 is a large multi-domain protein. It’s interesting that each of the domains except for the catalytic domain is needed for the condensation. In addition to a dsDNA that mimics double-strand breaks, condensation of PARP1 was also observed with nicked DNA, resembling single-strand breaks. What is the functional importance of this phenomenon? Nice optical tweezer-based experiments suggest that a PARP1 condensate prevents two sides of a double-strand DNA break from separating, plus the condensate could provide the initial repair environment. In cellular experiments, PARP1 seems to share with KU70 the role of preventing DNA ends from separating after a cut, the double loss of both proteins leading to the loss of bridging. PARP1 is also an enzyme, which gets activated upon DNA break binding and generates poly(ADP-ribose) (PAR) chains from NAD+. It looks like PARP1 is catylytically active in condensates and PAR chains can weaken or altogether prevent condensation. But then there are proteins, like FUS, that can come in to stabilise the condensates by interacting with PAR chains, and one can imagine a complex evolution of the initial condensate in cell on arrival of different proteins. Trapped PARP1 would be PARP1 in condensates that cannot mature beyond the initial stage. Overall, I really enjoyed it & it made me ask myself a lot of questions when reading. Protein condensation, which may or may not involve phase separation, has always seemed to me a somewhat esoteric phenomenon, and I know that it is controversial. But condensation mediated by specific domain:domain interactions is easy for a structural biologist to swallow, and it makes sense that multivalent molecules should be able to crosslink (or ‘percolate’) into large isotropic networks that manifest as spheres in vitro or foci in cells. Congratulations to Nagaraja Chappidi, Simon Alberti, and all other Authors.
Modelling domain:peptide interactions with AlphaFold2 (24/01/2024)
I like to think that, as a rule, at least one partner involved in a binary protein:protein interaction has to use a folded domain. There might be exceptions to this rule - biologically significant interactions between two regions that cannot be characterised as a folded domain - but I think they would be rare or perhaps not rare but difficult to detect and characterise with the tools that we use as they would probably be weak and serve only auxiliary roles in a system already stabilised by more canonical interaction. The reason I think that in more canonical interactions at least one of the partners tends to use a folded domain is that a domain can have a binding pocket with which it can at least partially surround a smaller ligand (such as a short linear protein motif) to achieve sufficient interaction surface and/or sufficient number of individual noncovalent bonds to get a meaningful interaction, which would be difficult if both partners were small. Now regarding the other partner, it might also be a folded domain or it could be just a short linear motif, typically embedded in a disordered region, which becomes more rigid upon binding to a domain. These kind of interactions, which are usually weak/moderate thermodynamically (in a micromolar range) and transient kinetically, could be described as domain:motif interactions. We know very many examples of such interactions and it’s possible that they are the most abundant type of binary protein:protein interactions, at least in higher organisms with complex interaction networks. In the SUMOylation system - by which I mean enzymes, substrates, and readers associated with the protein post-translational modification (PTM) called SUMOylation - interactions of this type are key and they include interactions between SUMO and the so-called SUMO-interacting motifs (which can be found in SUMO reader proteins and in SUMO E3 ligases), but also interactions between the SUMOylating E2 enzyme UBC9 and the so-called SUMOylation consensus motif on substrates. With the recent advances in AlphaFold2, many people have been asking if this tool can be used for predicting protein:protein interactions, including domain:motif type, and we also try using it to study these type of interactions in the SUMOylation system. Not being a computational biologist by training, I try to follow the literature by experts in the field, which by the way is moving fast. Recently there have been two papers out - both previously released as preprints - that deal specifically with the question of domain:motif interactions: this one and this one. They are partially overlapping but mostly complementary and I think it’s great to read both. I’ve finished the first and am halfway through the second. The first is by Hélène Bret as the first author and Jessica Andreani and Raphaël Guerois as supervisors, working not far from us in Gif-sur-Yvette near Paris. What is great in the paper is not just the results but also a very useful introduction and similarly useful discussion, in which the recent literature and the available methods and tricks are discussed in an accessible way. Indeed, AlphaFold2 seems to be an “art” as much as “science” and a lot is about finding the right protocols to run it most efficiently. I noted down a few papers with which I have to catch up. The bulk of the results presented in the paper is based on a set of 42 interactions between a structured domain and a short motif embedded in a disordered region. This set comprises new structures not included in the training set of AlphaFold2-Multimer and also lacking homologues in that set. This set is then used to evaluate how good AlphaFold2-Multimer, applied in different ways (with full-length proteins or fragments, with or without multiple-sequence alignment, with different types of alignment, etc.) is at predicting such interactions. One interesting conclusion - which aligns with our experience and probably those of some others, but we haven’t analysed it systematically yet - is that prediction works better if sequences are limited to relatively short fragments spanning the interaction regions (ideally not more than 100 aa on the motif side; for the domain, one can, and probably usually should, take the whole delimited domain). One can scan the disordered sequence with such windows to find the motif if its location is unknown. A multiple-sequence alignment (MSA) of the disordered fragment is particularly beneficial if long sequences are used, but if sequences are delimited, it makes surprisingly little difference and just having the MSA of the domain is enough. (Sometimes including an MSA of the disordered fragment might even be detrimental, as when it makes AlphaFold2 fold the interacting motif in a structure that is not compatible with binding! - there are some interesting examples of that discussed in the paper). In the words of the authors, “Such a performance suggests that the structural and evolutionary properties of the receptor match well with the peptide sequence irrespective of its conservation pattern”. As far as I understand, there is a lot of discussion in the field which metrics it is best to use to rank AlphaFold2 models and identify potentially real ones. In this paper, they use a linear combination of 0.8 * ipTM + 0.2 * pTM, which seems to work well, so it might be a useful, simple metric to use. Thanks to the authors of these lovely papers!
Review article, article review (21/12/2023)
I’ve finished revising and re-submitted a general review on chemistry, mechanisms, and evolution of protein post-translational modifications (PTMs). This brought back the memories from 12 years ago or so, when I had just finished my undergraduate studies and did a 6-month internship at the Weizmann Institute in Israel, in the group of Joel Sussman.
It was a great, formative time for me. I shadowed a PhD student and did some enzyme kinetics measurements and crystallisation with him, learned to write scripts in Perl, and - which is why I started thinking about it again - wrote a review article - the very first scientific article of any type in my life. The topic was proteolytic stability of intrinsically disordered proteins in vitro and in cells. It started with Joel asking me for my opinion on a paper he had to review. Nobody had ever given me such a responsible task, so I approached it (too) ambitiously, read widely around, and wrote a long (too long) opinion on the article. Joel suggested we transform it from a review of a paper into a review article on the topic that the paper was about. I had then recently learned how to make simple schematics in a freeware equivalent of Illustrator called Inkscape, so I tried to make use of it for illustrating my little review. I don’t know if what we wrote there still remains up to date after 12 years, but I have fond memories of writing it, discussing it with supervisors and getting their feedback, and then of seeing it published. Since then, I have participating in writing a few more review articles, but the one now is special - and closest to the one 12 years ago - in that I did it mostly by myself and put extra effort. Let’s see what comes out of it. PTMs have been an interest of mine for many years, the main unifying theme that connects different projects I have been doing over the last 12 years or so, since my undergraduate studies and that internship with Joel. Studying different particular PTMs, first in bacteria (protein arginine phosphorylation) and then in eukaryotes (ADP-ribosylation, ubiquitylation, SUMOylation…), I started asking myself questions about the chemical nature of these processes, and how it is connected to, on the one hand, their catalysis, and, on the other hand, their emergence and development during evolution. In short, what I think is interesting is that most major PTMs are chemically analogous in that they all involve some kind of nucleophilic substitution reaction where a protein reacts with a donor molecule present in the cell (usually present there for other reasons, and present already before PTM reactions emerged - examples of such donors include ATP, NAD, SAM, acyl coenzymes A, UDP-sugars, etc., all of which are involved in metabolic processes in addition to protein modification). In these donor molecules, one can distinguish a good leaving group, which leaves during the reaction, the rest of the donor ending up covalently attaching to the protein. So enzymes that catalyse PTMs tend to perform similar tasks, binding the donor in such a way that it promotes the departure of the leaving group and the nucleophilic attack, and often they deprotonate an amino-acid residue on the protein to make it more nucleophilic and attack the donor more efficiently. In the review, I try describing these chemical and catalytic analogies, but also analogies between different PTM systems in terms of how the substrates are recognised by the writer enzymes, how the modification might be reversed by eraser enzymes, and how the product is recognised by reader proteins. And, lastly, I try to discuss the evolutionary emergence of these processes, where one part of the story is that the suitable “activated” donor molecules had existed in primordial prokaryotes to serve in primary metabolism and then became repurposed as co-substrates of enzymatic protein modifications. That’s a very brief summary of the review, which will perhaps be out in a few months. There will be some figures, too. Now I have a license for the real Adobe Illustrator, but I still make the same sort of simple schematics to illustrate the processes I write about.
So that’s one thing I’ve been doing in the last few days. I also had to do some things in the wet lab (mutagenesis and some recombinant protein production, autoclaving media, etc.), and today I wrote a review of an article. Since very recently, I am (very sporadically) asked to do it directly by journals, which is flattering but also some responsibility. With the review article and the article review now submitted and this blog post finished, tomorrow I start two weeks of holidays for Christmas.
Paper on PARP4 (01/12/2023)
I’ve finally read this study of PARP4 by Léonie Frigon and John Pascal and strongly recommend it to those interested in PARPs and ADP-ribosylation (one of my interests, especially during my ‘previous life’ as a postdoc). PARP4 is an odd cousin of the canonical PARP protein, PARP1, sharing a larger homology region with it than any other of 17 or so PARP family members in humans. At the same time, PARP4 is quite diverged from PARP1 in terms of cellular location and function. Previous work, largely from the Pascal lab, showed that PARP1 is an amazing allosteric machine that is very tightly regulated. Now PARP4 can be seen as an example of evolution duplicating and rewiring this molecular machine, keeping some functions and chaning or modulating others.
I enjoyed the thoughtfully crafted and cautious text that leaves room for discussion and interpretation of results, going through various important aspects of PARP4 and how it seems to differ from PARP1, including the regulation by HD, the absence of the PAR chain extension function in PARP4, the potential roles of WGR and BRCT domains. For example, it looks like the HD of PARP4 does not inhibit its catalytic activity much (unlike in PARP1), and the WGR apparently lost its nucleic acid-binding function, while BRCT retains it, possibly prefering vault RNA over other nucleic acids, although that is not completely clear.
Earlier this year we published a paper that analyses AlphaFold2 models of all PARP family members, including PARP4. I’m sorry if it has in some ways impinged on the apparent novelty of this article by Frigon and Pascal or caused any stress with their publication. I don’t think it did impinge on the novelty. Indeed, we only now get a real experimental analysis of the N-terminal catalytic region of PARP4, combining crystallography with functional in-vitro assays.
Working at the bench (27/11/2023)
I do hope I will be able to continue working with my own hands at the bench still for some time, rather than focussing only on administrative and supervision roles. I do enjoy the wet lab a lot and also think working with your own hands is useful for thinking about projects. There are many great group leaders who do not work at the bench any more themselves, but I would like to do it if I can. I’ve been in a meeting with a Nobel prizewinner for ubiquitin discovery, Avram Hershko, once, and he said that even now, after the big prize etc., and in his wisdom years, he works at the bench, usually from the early morning until around midday, where he begins to take care of other things. That sounded great. On this topic, a quotation I’ve found, about the pioneer of fly genetics, Thomas Hunt Morgan: “Throughout their careers Morgan and students worked at the bench. The investigator must be on top of the research if he or she is to recognize unexpected findings when they occur.”
At home and away (27/11/2023)
I do enjoy travelling, although much less since I have a family, unless, of course, we can go together. The last two months or so have been a period of several work-related trips, mostly short and nearby (all by train), but still amounting to quite some time away from home and the lab, and only one of the trips was together with my family.
One, alone, was to a local conference organised mainly by and for scientists from Tours and Orléans, the yearly Colloque Biotechnocentre. It was its 35th edition already. The conference is held in French and takes place on two consecutive days, with an evening (with a party) in between. There are poster sessions for PhD students and talks by invited guests from outside the region and local scientists, both more senior and PhDs. I had a short talk entitled ‘Why we study protein SUMOylation?’, or rather ‘Pourquoi étudier la SUMOylation des protéines ?’, because it was the first proper scientific talk that I’ve given in French, with much fear and trembling. But it was a nice experience in the end, having managed to do it and seeing that I have been mostly understandable. I enjoyed this particular yearly conference very much. I like its broad scope in terms of topics but at the same time the regional focus, and I like the multi-generational make-up, with both older (some retired for some time) and young scientists, and last but not least the fact that it combines an intensive scientific programme with a party and long French-style meals etc. At the same time, it is relatively modest in terms of venue etc. and not too expensive. I was very impressed by PhD students - their talks and posters were very well prepared and well presented and the atmosphere was friendly. By the way, I had a very similar impression at a PhD defence I attended in Rennes as a member of a jury in early November - of being impressed by the PhD students, both the one defending (she did it great), but also those attending the small party afterwards, being there for her.
Another recent occassion was a meeting of prize-winners of the French ATIP-Avenir programme. A great meeting as well, and what I particularly liked this time was that they invited some older scientists (some of them themselves ATIP-Avenir laureates from the past) to give talks that were not pure scientific talks but rather attempts to share experience of running a group and developing your research after the first grant. Two of the speakers whose names stayed in my head were François Parcy and Jean-Louis Mandel, but there were many others. There were also short talks by the ATIP-Avenir awardees themselves, including myself, presenting the selected projects. I should say that because of the rules of the ATIP/Avenir programme, getting an ERC grant means you don’t get any money from ATIP - but you still get a ‘label’ of an ATIP-Avenir programme and get to attend the events like this, which is nice (there were several people in this situation).
Finally, the last two trips - 5 days in Marseille on the biannual French structural biology meeting called BSI, and then a seminar in Basel, at the FMI institute, last Friday. But let me write more about the BSI conference. It was the second BSI meeting I have attended in my life, after the one two years ago in Saclay, which was one of the first things I did after starting the job in France. The BSI meetings are really good. Quite diverse in terms of biological questions and techniques (ranging from EM and crystallography to more niche biophysical techniques such as AFM or EPR), with a diverse set of speakers at different career stages (most of them from France, but there are always some invited speakers from other countries). It is a good opportunity to get a sense of how the field is developing, at least in France (e.g. this year - the growing interest in EM tomography and, obviously, in AlphaFold). I had a talk in which I presented, for the first time, a new topic in the lab. The talk went ok, I think, but I didn’t have it finished before the conference so the evenings before my talk were stressful and busy with rehearsals. There was a lovely ‘gala dinner’ during the conference in a museum, on the last floor, overlooking the harbour of Marseille.
It was at the BSI in Marseille that I had a conversation with one young scientist about science as a social phenomenon. This social dimension of science - the invisible hierarchies and codes that govern it - has always fascinated me. Conferences or visits at other labs are great opportunities not only to learn about new discoveries or establish collaborations - although they are both - but also to look at science for a moment with an outsider’s eye, which allows noticing various peculiarities of how sience works.
But there is also a price to travelling, especially being separated from one’s family and putting an unequal burden on the spouse/partner. I hope I will travel less in the future.
Nobels 2023 (06/10/2023)
Like every year, I’d been eagerly awaiting the announcement of Nobel prizes. Like everybody else, I was very happy to hear that Katalin Karikó and Drew Weissman were awarded, especially Karikó, but what I think made me even happier was the Nobel prize in literature awarded to Jon Fosse, a Norwegian writer whose thick Sepatalogy I began reading some time ago and which has enchanted me in a special way. A running joke is that Fosse’s new novel is easy to read, because you just need to read one sentence (there are no full stops in the whole long book). Well, easy it is not, but it is a particularly convincing example of a stream-of-consciousness narrative, and the writing has a singular trance-inducing rythm.
Grenoble revisited (04/10/2023)
We’re going to Grenoble today, just me and a colleague, for a couple of days, to do some SAXS experiments at the synchrotron. I think it’s been about 12 years since I first visited the ESRF synchrotron. It was before my PhD, during an internship I did in the group of Joel L. Sussman at the Weizmann Institute in Israel. We spent a few days in Grenoble, collecting data for two days or so and also doing a little bit of sight-seeing (we took a cable car to a base beneath Mont Blanc and stopped in Chambéry). I remember the Hebrew word “maruah” that we used when poorly diffracting crystals gave a smeary pattern. In addition to it being the first time at a synchrotron, it was one of the first encounters with France. Eating croissants in the ESRF guesthouse for breakfast, little did I know that I would one day live and work in France. I’ve been to Grenoble a few times since, but not in the last 5.5 years or so, so I’m happy to go back. And it’s the first time I’ll do SAXS there.
Added on 6th October: So now we’re actually in Grenoble, and almost finished and leaving very soon. It’s been a lovely couple of days. We’ve met some kind people at the beamline and outside of it, including some old friends and a postdoc behind the data of one of my favourite talks in the Aviesan meeting mentioned below. The scenery of the Polygone Scientifique of Grenoble, with mountains all around, and the atmosphere - all the scientists, including many young people, working here or just coming to collect data - it’s all very special.
Aviesan ‘Membrane-less organelles and liquid condensates’ symposium, Paris 25th September 2023 (26/09/2023)
I very much enjoyed yesterday’s one-day symposium on membrane-less organelles organised by Aviesan, one of those enigmatic French organisations that I haven’t figured out yet. What I’ve figured out so far is that it is an alliance of scientists that is reputed for organising very nice free symposia on different topics, and this was one of these. The main organisers of this specific symposium were Carine Giovannangeli, Yves Gaudin, and Dominique Weil. The symposium was held in Jacques Monod Institut in Paris (my first time there), so very conveniently near the Austerlitz Station where trains from Orléans arrive. I don’t know the field of biomolecular condensates very well and have been at times sceptical of some very general claims that I heard, but the symposium made me realise that there is a lot of nuance in the field, like, for example, an attempt to distinguish polymer polymer phase separation or mere binding-driven phenomena from (perhaps rarer?) liquid-liquid phase separation, or the realisation that ‘membrane-less organeless’ often result at least in part from defined and specific (even if weak) binding events, or the idea that there is sometimes some degree of order within these apparently liquid structures. So there is nuance and more theoretical and technical innovation, but there is also certain dynamism and audacity characteristic of new fields and new paradigms, and I enjoyed that aspect a lot. I think my favourite talks were those by Chloe Zubieta, Ariel Lindner, Fabian Erdel, Martin Blackledge, but I liked all of the others too, including by young scientists, which did a very good job (a postdoc Damien Glon, for example, and Anastasiia Skolbelikna). Overall, a great line-up of talks spanning a bit the whole field in France, with both some finished studies and some work in progress. The main keynote lecture by Anthony Hyman - the only speaker outside of France - was on Zoom but the technology worked well. I’m looking forward to more events like this.
Protein homodimerisation (20/09/2023)
A lot of proteins we see in nature are homodimers in which the dimerisation interface is formed by two identical surfaces, one on each subunit, coming together. This is how many proteins have evolved to be, and various functional advantages of homodimerisation (things like cooperativity, or higher stability, or easier folding, etc.) might have driven this evolution. But, generally speaking, for a feature to be selected for and optimised over time, it has to already exist in a primordial system, and perhaps if there is a bias towards that feature already at the start, it can be more easily picked up and optimised over time. Is there, therefore, an inherent bias for a random protein surface to associate with itself rather than with a different protein surface? This is a question posed in this somewhat old paper from Lukatsky et al. (the Shakhnovich lab). The paper contains a simplified model of a protein surface through which this question is investigated computationally (giving an affirmative answer), but there is also a common-sense argument, which I like, and which I think can be rephrased in the following way. What is special about a typical homodimerisation interface? That it has what crystallographers call 2-fold rotational symmetry (C2 symmetry), which means that all (or most) interactions are repeated twice. If residue X from subunit 1 interacts with residue Y from subunit 2, then residue Y from subunit 1 interacts with residue X from subunit 2. This repetition has implications for the likelihood of making homodimers, because, if you make certain favourable interactions by chance, they will be repeated twice. I think this is similar to what Lukatsky et al. say here: “it is more probable to symmetrically match a half of a random pattern with itself than with a different random pattern, which requires a full match”. If we imagine that dimerisation is pattern-matching, it is more probable to match half of a pattern (& symmetry does the other half) than to match a full pattern (which is what has to happen during hetero-dimerisation)
I think that the repeptition idea also implies that the rate with which an interface becomes optimised over time is higher for a homodimer than a heterodimer - because a favourable mutation that strengthens the interface would have an amplified effect (would effectively be a double mutation). As Monod and his colleagues working on haemoglobin already observed, symmetry amplifies effects. Perhaps this is the common-sense rationale behind another modelling study, this time from G. E. Schulz and with a more sophisticated model tracking evolution over time. This second study concluded that a simulated evolutionary process that leads to homodimerisation has the probability that is by a factor of 100 or above higher than the probability of an analogous simulated process that leads to heterodimerisation.